Indomyrma
| Indomyrma | |
|---|---|

| |
| Indomyrma dasypyx | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Arthropoda |
| Class: | Insecta |
| Order: | Hymenoptera |
| Family: | Formicidae |
| Subfamily: | Myrmicinae |
| Tribe: | Crematogastrini |
| Alliance: | Vollenhovia genus group |
| Genus: | Indomyrma Brown, 1986 |
| Type species | |
| Indomyrma dasypyx | |
| Diversity | |
| 2 species (Species Checklist, Species by Country)
| |


This small Asian genus is known from only two species and is rarely encountered. Workers have been collected in forest habitats and both species are believed to nest in rotten wood.
Photo Gallery

Identification
General (2015) - Indomyrma has reduced eyes with a total of less than 12 ommatidia, an antennal scrobe, albeit weakly developed, bounded dorsally by a distinct frontal carina, median clypeal margin expanded as a notched convex apron that barely covers basal borders of completely closed mandibles, flat dorsum of promesonotum and propodeum, erect hairs in bilateral positions on head, mesosoma, petiole and pospetiole, propodeal spines present, and a pedunculated petiole (Brown 1986).
Brown (1986) - Worker: Monomorphic myrmicine ant with 11-merous antennae and 3-merous antenna] club, similar to Dacetinops, but with the following principal differences.
(1) Spongiform appendages completely lacking (on petiole, post petiole and base of gaster).
(2) Petiole nodiform, with distinct anterior peduncle.
(3) Antennal scrobes shallow and weakly demarcated, open to dorsal view.
(4) Frontal lobes smaller and more mesally positioned, only narrowly separated.
(5) Basal tergum of gaster (abdominal tergum IV), which takes up most of gaster, shallowly and closely, but distinctly foveolate throughout, with minute roughening between foveolae, opaque, basal longitudinal costulae not apparent.
Queen: Alate when virgin, differing from the worker in slightly larger size, much larger eyes, and the usual differences seen between female castes in the subfamily. Wing venation of the Tetramorium pattern, with forewing RS not reaching costa (“radial cell open”); hind wing narrow and with sharply rounded apex.
Male: typical myrmicine male, smaller than queen, with similar wing venation, but with aberrant 12-merous antennae having a strongly S-shaped curvature or kink near and beyond midlength, caused in large part by a deformity of antennomere VII, which has its basal half greatly narrowed and somewhat curved.
Genitalia essentially typical of small-sized “core” Myrmicinae, except for aedeagus, which is short and broad and has a deeply emarginate (bilobed) apex.
Larva: similar to that of Dacetinops (allowing for instar differences and small sample size of the material available).
| See images of species within this genus |
Distribution
Distribution and Richness based on AntMaps
Species by Region
Number of species within biogeographic regions, along with the total number of species for each region.
| Afrotropical Region | Australasian Region | Indo-Australian Region | Malagasy Region | Nearctic Region | Neotropical Region | Oriental Region | Palaearctic Region | |
|---|---|---|---|---|---|---|---|---|
| Species | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 |
| Total Species | 2841 | 1736 | 3045 | 932 | 835 | 4379 | 1741 | 2862 |
Biology
See the individual species pages for notes regarding the biology of the two species of Indomyrma.
Castes
Queen

Head view

Side view


Top view

Specimen labels





Male

Head view

Side view

Top view

Specimen labels




Morphology
Worker Morphology
 Explore: Show all Worker Morphology data or Search these data. See also a list of all data tables or learn how data is managed.
Explore: Show all Worker Morphology data or Search these data. See also a list of all data tables or learn how data is managed.
Male Morphology
- Explore: Show all Male Morphology data or Search these data. See also a list of all data tables or learn how data is managed.
• Antennal segment count 12 • Antennal club 0 • Palp formula 2,2 • Total dental count 1 • Spur formula 0, 0
Phylogeny
| Myrmicinae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
See Phylogeny of Myrmicinae for details.
Nomenclature
The following information is derived from Barry Bolton's Online Catalogue of the Ants of the World.
- INDOMYRMA [Myrmicinae: Stenammini]
- Indomyrma Brown, 1986: 37. Type-species: Indomyrma dasypyx, by original designation.
Unless otherwise noted the text for the remainder of this section is reported from the publication that includes the original description.
Description
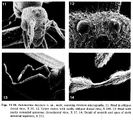
- Brown 1986. Figures 1-21









{kind=link}
Worker
Head broad behind, narrowed anteriad, with convex sides in full-face view; posterior outline weakly concave in middle third, the posterolateral corners more rounded seen in dorsal view than in Dacetinops, but still with a narrow flange or “ear” as seen from side view. Eyes small but convex, with less than 12 (usually 8-10) distinct ommatidia (Fig. 5), situated a little anterior to midlength of head, ventral to ventral limit of scrobe.
Frontal lobes well developed, convex, and with rounded sides, but situated close together toward median line of head, separated by narrow, impressed, posteromedian extension of clypeus with adjacent, sunken, long-oval shining frontal area, which together form a keyhole-shaped frontal cleft between lobes. Lobes pinched in slightly caudad, but continued as fairly distinct but obtuse frontal carinae, that diverge gently toward posterior corners of head where they fade out at about posterior tenth of HL after forming dorsal margins of shallow scrobes much less distinct than those of Dacetinops. Frontal lobes rounded anteriad and situated well back from anterior margin of clypeus. Clypeal shield narrower than head across mandibular insertions, but long anteroposteriorly; median portion just in front of frontal lobes convex, but falling off steeply to sides and dropping anteriad to a narrow apron or marginal lamina that is strongly convex in outline, but with a small median notch (Figs. 1 ,3). Clypeal apron barely covers basal borders of fully closed mandibles. Raised median portion of clypeus not clearly carinate, but its lateral margins fall off so steeply on each side that the angles or edges so formed, diverging in wide curves around the anterior margins of the antennal fossae, may be taken as the homologs of the twin divergent carinae in other myrmicine genera (e.g., Rogeria ), and perhaps even of the antefossal ridges characterizing Tetramoriini and some primitive Myrmica species. It should be recalled that similar carinae bound the median clypeallobe of ectatommine ponerines such as certain Proceratium.
Mandibles robust, triangular, closing together fully, masticatory borders straight, overlapping slightly apicad when closed; bases narrowed and gently incurved; basal borders gently convex, distinct from masticatory borders, hidden beneath clypeal apron at full closure; masticatory borders each with 9 (8-10) separate teeth, the apical tooth largest and sharpest, followed basad by 2 teeth decreasing in size, then by 6 (5-7) irregular denticles occupying the rest of the border. Lateral mandibular border, as seen in full-face view of head, convex at base, straight to very feebly concave in the middle half of its course, then convex to end of apical tooth.
Labrum wider than long, apical margin broadly emarginate (or bilobed), extensor surface weakly convex, shining. Palpi short, segmented 2, 2.
Antennae with stout, weakly sigmoidal scapes that broaden gradually from base to apex, with 11 segments total; pedicel (II) about equal to III+IV+V; III-VI broader than long; VII and VIII slightly longer than broad; IX, X and XI much longer than broader, together forming a club longer than remainder of funiculus. Club 3-merous; the preceding segment is slightly enlarged, and could be considered as a fourth segment of the club, but this would be stretching a point.
Alitrunk well represented by figs. 2, 6 and 9, so the description here will be limited to emphasis on important features and clarification of those that may be obscure in the figures. First to be noted is the compact, sutureless dorsal surface, evenly convex from the distinct anterodorsal pronotal margin (flanked by subacute humeral angles) to the shallow but distinct metanotal groove, which is not clearly shown in the photographs. Behind metanotal groove, anterior half of propodeal dorsum convex, after which posterior half becomes concave onto bases of upwardly inclined propodeal teeth, which themselves have prevailingly upcurved dorsal margins and apices, so that dorsal propodeal outline as seen from the side is distinctly sigmoidal. These features, as well as the strong, oblique pleural groove indicating meso-metathoracic suture, are somewhat obscure in the figures. Propodeal spiracles prominent, situated very close to declivitous margins, directly below tooth bases, their openings facing almost as much caudad as laterad.
Metapleural gland bulla moderately distinct, meatus small and obscure, but with two curved, longitudinal sulci leading forward along lower metapleuron (Fig. 2). Propodeal declivity distinct, higher than wide, margined above by one or two curved, transverse costae connecting the two teeth, and by thin, sharp lateral margins that merge into the metapleurallobes (inferior propodeal plates, fig. 2); declivitous surface weakly shining, very finely reticulate-punctulate, becoming smooth and shining ventrad.
Petiole with a short but distinct anterior peduncle and a subcubical node; peduncle broadening and thickening caudad to meet node {Figs. 3, 6) and obscurely marginate above on each side; anteroventrad, the anterior peduncle bears a distinct, weakly curved, digitiform, subpetiolar process directed obliquely cephalad between coxae, and visible when gaster with the two waist segments is raised slightly. Petiolar node highest near anterior angle, then falling slightly through a convex surface to a distinct but narrow posterior collar. Viewed from above, petiolar node nearly square, about as wide behind as it is long, but very slightly narrower cephalad; anterior margin straight to feebly convex, depending on exact angle of view, and anterior corners almost rectangular in direct dorsal view, which is not the view in fig. 6. A minor feature is the presence of a pair of shallow, fine, converging sulci on posterodorsal surface of node, not visible in figures.
Post petiole shorter than petiole, but distinctly broader, and broader than its own length; dorsum and sides strongly rounded.
Gaster broader than deep (about 7:5), and basal tergum (true T4) covering almost 0.9 of its length in dorsal view. From dorsal view also, middle section of anterior margin straight, or even feebly concave, so that basal outline appears slightly shouldered on each side. Lateral suture not arched basad, running well below bulging overhang of tergum along sides of gaster. Apical segments (A7 and succeeding ones) forming a small, blunt cone, directed slightly downward as well as caudad.
Sting (worker and queen) not dissected, described only from dried specimens with shaft and part of sting bulb, plus gonostyli, exposed. Sting shaft long and slender, almost straight distad of the slightly arched bulb, almost imperceptibly upturned apicad, the visible parts resembling figs. 75, 76 (Vollenhovia), fig. 92 (Rogeria) or figs. 146, 147 (Stenamma) in Kugler 1978. Gonostylus long and gradually tapered to an acute apex; probably bipartite, although the articulation, if it exists, is nearer to the base than the apex. It will be interesting to see the results of a properly detailed study of the sting structures.
Legs unremarkable; femora and tibiae robust, hind tibia nearly 2 X as wide as its thickest point as its basitarsus; tibiae II and III spurless at apices. Tarsomeres 11-V taken together slightly shorter than I, and each taken separately slightly longer than broad. Tarsal claws slender, simple; arolium undeveloped, replaced by a pair of short, slender, erect hairs.
Sculpture distinctive, opaque or neary so over most of body, consisting mainly of distinct, round, centrally tuberculate and piligerous foveolae (Figs. 1, 2, 3, 6, 9), crowded and subcontiguous on both dorsal and ventral surfaces of head, especially on anterior and lateral aspects, on dorsum of alitrunk, nodes and gastric tergum I (more shallow and with partly shining interspaces here apicad). On dorsum of head, foveolar interspaces form 8-9 slighly vermiculate longitudinal costae, including frontal carinae, between antennal scrobes; middle costae very distinct. Elsewhere, foveolar interspaces, alitruncal pleura, antennae, legs and sides of nodes finely and densely puctulate to reticulopunctulate. Gastric interspaces finely rugulose. Mandibles smooth and shining, with scattered fine punctures.
Pilosity consisting of widely distributed, moderately abundant, short, decumbent and subdecumbent setae; this vestiture denser, finer, shorter and more nearly appressed on antennae and legs. Setae fine and sparse on clypeus and mandibles. Longer, fine, curved, erect and suberect hairs, mostly 0.05-0.18 mm long, tend to arise from constant, bilateral positions on head, alitrunk and nodes: 2 pairs from median clypeal shield, 1 pair from frontal lobes, 3 pairs from on or just mesad of frontal carinae at different distances from front to rear, 1 pair (often lost) from near posterior corners of head, a humeral pair, plus about 6 more pairs on pronotal disc and mesonotal dorsum; 2 pairs suberect marking dorsal corners of petiolar node, 3 pairs on postpetiolar tergum, inclined caudad. Gastric tergum with many of these longer hairs, curved sharply caudad, and gastric sternum with similar but shorter hairs.
Color brownish red; appendages and mandibles dull yellowish; some specimens darker, mahogany approaching black, and appearing black to naked eye.
Male
Alate and large-eyed, of usual myrmicine habitus, except for antennae, which are 12-merous and aberrant, as described below and shown in figs. 13 and 14. Head without eyes slightly longer than broad, broad behind and broadly rounded in dorsal-view outline; tapered anteriad in front of eyes, convergent sides continued in outline by the closed, narrowly sub triangular, converging mandibles, opposable and with basal margins convex and broadly continuous with convex masticatory margins, which are cultrate and end in a small, acute tooth at each apex. Mandibular shafts bent slightly mesad near their insertions.
Frontal lobes small (Fig. 11 ); frontal area small and indistinct; a shallow median frontal line or sulcus leads back toward median ocellus; scrobes not developed. Clypeus convex, broadly triangular, narrowed posteriad and inserted between frontal lobes, but broadly and very weakly convex apicad. Labrum broad, with a weakly convex, shining dorsal surface and bilobed at apex, found extended in most specimens, filling most of the otherwise empty space between closed mandibles.
Under-mouthparts not dissected in male, but the palpi are short, apparently dimerous and similar to those of the female castes. ,
Antennae (Fig. 13) slender, contorted in about middle third of length, antennomeres mainly cylindrical basad, III long, subequal to IV-IV together. Apical segment (XII) long, fusiform, slightly incrassate; antennomere VII (Fig. 14) of a peculiar shape, with very slender, curved, stalked portion basad, widening to a more normal thickness at apex. This and the more normally formed, but curved, succeeding (VIII-XI) antennomeres together form a semirigid, more or less S-shaped mid section of the antennae, including the sharply narrowed VII, the function of which is seen in the living colony to serve as a handle by means of which the passive, folded male adult or pupa (pharate adult) is carried in the mandibles of workers. This structure of the antennae is uniform as seen in all males in at least two different colony series from different localities, and it appears to be naturally preformed, in the course of development, not just the secondary result of pressure from worker mandibles.
Pronotum inverted U-shaped around the inflated mesonotum, which bears distinct, impressed, convergent notauli that almost, but do not quite, meet in a V cuadad. Parapsidal furrows present as fine impressed lines. Scutellum and the much smaller metanotum both distinct, convex, and bulging, but only modestly so. Propodeum with steeply sloping dorsal surface nearly twice as long as declivity, and nearly flat, separated from declivity by obtuse angles representing propodeal teeth; declivity prevailingly weakly concave in both lateral and dorsal views, with blunt, submarginate lateral boundaries ending ventrad in rounded inferior propodeal plates or flanges. Propodeal spiracle near the middle of the lateral propodeal face, well away from declivity.
Petiole long and low; node with low, gently sloping anterior and posterior surfaces, and thus indistinct in side view, but in dorsal view, node broader than long. Subpetiolar process represented in the male only by a minute, subangular terminus of the median sternal carina. Postpetiole only about half as long as petiole, but slightly wider, with strongly rounded dorsum and sides of node; briefly pedunculate cephalad, and broader, but sharply constricted caudad.
Gaster broader than high, without shoulders basad, and sides evenly convex throughout; basal tergum constituting nearly 3/4 of total gastric length. Legs slender, unremarkable, the mid and hind tibiae spurless; tarsal claws slender, weakly curved; arolia absent, replaced on each pretarsus by a pair of fine, short, erect hairs. Basitarsus long and slender, especially on hind leg (average L about 0.55 mm, hind tibia L 0.50 mm).
Wings like those of queen, but a little smaller (average forewing L 2.7 mm) and with discoidal ceil more nearly triangular (narrower at anterior apex, due to shorter RS + M); veins brownish, and membrane sown densely with minute, brown microtrichia. Hind wing narrow; costal margin with 4 submarginal hamuli, as in queen.
Genitalia completely retractile, the dissected capsule shown in figs. 15-18. Distinctive feature is the deeply notched ventral margin of the paramere (on right side in fig. 18, and especially the short, broad, apically emarginate aedeagus (Fig. 16)). The paramere also has some sort of mesal process near its apical quarter, but the details are still unclear (Fig. 17).
Except for the smooth and shining (micropunctulate) gaster and labrum, body and appendage sculpture prevailingly a very fine, shallow, matt, reticulo-punctation, passing into peculiar, “deep” striolation on vertex (Fig. 12), and variably overlain by fine, spaced costulation on propodeum - vertical on sides, tranverse above on anterior dorsum.
Most of dorsal body surfaces, appendages, and undersides of head and gaster covered by copious, short, fine, appressed to decumbent, but mostly inconspicuous pubescence. Longer, but still mostly short, fine, sparsely arranged hairs on clypeus, vertex, underside of head, dorsal surfaces of pronotum and pterothorax, paired on sides of petiole and postpetiole and postpetiolar tergum, and gastric apex.
Color brown, lighter on alitruncal pleura, mandibles and appendages, darker above on alitrunk and gaster; head very dark brown or piceous to nearly black.
Larva
Mature worker larva: L through spiracles about 2.2 mm, straightline L 2.0 mm. Profile aphaenogastroid (G.C. and J. Wheeler 1976: 76), but the body constriction weakly marked. Considering that this is a mature larva, and that the larva previously described for Dacetinops (G .E. and J. Wheeler 1957) was only a small one of an earlier instar, the correspondences between them in head and body form, mouthparts, and pilosity are very close. I was not able to distinguish the antennae in my preparations, and I am not sure about the number and types of sensilla present on the cephalic appendages, but in other characters discussed by the Wheelers, the similarity between lndomyrma and Dacetinops cibdela larvae is nearly complete. I would mention as minor differences that the mandibles of Indomyrma are slightly stouter, especially basad, and that there are a few reclinate spinules on their inner (mesal) surfaces on the basal half. The bilobed labrum in Indomyrma is 3.5X as broad as long. Some of the head hairs, bifid at the apex, are seen under magnifications of 400X or more to be finely denticulate or twigged within the split apices. The ventral welts of abdominal somites II and III in Dacetinops are much less pronounced in these older Indomyrma larvae. The anchor-tipped hairs in transverse rows of 4 each across the dorsal somites are longer and have kinkier basal shafts that in the young Dacetinops larva. On the basis of the evidence available, I do not see how Dacetinops and Indomyrma can be separated on larval characters.
References
- Blaimer, B.B., Ward, P.S., Schultz, T.R., Fisher, B.L., Brady, S.G. 2018. Paleotropical diversification dominates the evolution of the hyperdiverse ant tribe Crematogastrini (Hymenoptera: Formicidae). Insect Systematics and Diversity 2(5): 3; 1-14 (doi:10.1093/isd/ixy013).
- Bolton, B. 1994. Identification guide to the ant genera of the world. Cambridge, Mass.: Harvard University Press, 222 pp. (page 106, Indomyrma in Myrmicinae, Stenammini)
- Bolton, B. 1995a. A taxonomic and zoogeographical census of the extant ant taxa (Hymenoptera: Formicidae). J. Nat. Hist. 2 29: 1037-1056 (page 1050, Indomyrma in Myrmicinae, Stenammini)
- Bolton, B. 1995b. A new general catalogue of the ants of the world. Cambridge, Mass.: Harvard University Press, 504 pp. (page 217, Indomyrma in Myrmicinae, Stenammini)
- Bolton, B. 2003. Synopsis and Classification of Formicidae. Mem. Am. Entomol. Inst. 71: 370pp (page 203, Indomyrma in Myrmicinae, Stenammini)
- Brown, W. L., Jr. 1986 [1985]. Indomyrma dasypyx, new genus and species, a myrmicine ant from peninsular India (Hymenoptera: Formicidae). Isr. J. Entomol. 19: 37-49. (page 37, Indomyrma as genus)
- Cantone S. 2018. Winged Ants, The queen. Dichotomous key to genera of winged female ants in the World. The Wings of Ants: morphological and systematic relationships (self-published).
- Zryanin, V.A. 2012. A new species of the genus Indomyrma Brown, 1986 (Hymenoptera: Formicidae: Myrmicinae) from Vietnam. Russian Entomological Journal 21:23-228.