Myrmica rubra
| Myrmica rubra | |
|---|---|

| |
| Myrmica rubra | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Arthropoda |
| Class: | Insecta |
| Order: | Hymenoptera |
| Family: | Formicidae |
| Subfamily: | Myrmicinae |
| Tribe: | Myrmicini |
| Genus: | Myrmica |
| Species: | M. rubra |
| Binomial name | |
| Myrmica rubra (Linnaeus, 1758)
| |
| Subspecies | |
| |
| Synonyms | |
| |


| Common Name | |
|---|---|
| Kiiro-kushike-ari | |
| Language: | Japanese |
A widely distributed, common European species that has been introduced in the northeastern and northwestern United States and western Canada. It is unusual in being an invasive species in temperate habitats; most ants that have spread by human activities occur and are spread in tropical and subtropical areas. In Japan, it nests in the soil of grassland on seashores and lowlands (Onoyama, 1989); rarely encountered (Japanese Ant Image Database).
| At a Glance | • Invasive • Polygynous • Supercolonies |
Photo Gallery
_(Tom_Murray).JPG)
_(Tom_Murray).JPG)


Identification
A member of the rubra group. Yellowish brown. Sculpture dilute; frontal triangle and subspinal areas smooth and shining. Antennal scapes long and slender. Petiole node with short indistinct dorsal area sloping evenly without definite break to its junction with the postpetiole. Head Index: 79.5; Frons Index: 49.4; Frontal Laminae Index: 92.7. Length: 3.5-5.0 mm. (Collingwood 1979)
Japan
This species closely resembles Myrmica kotokui, and is at times difficult to distinguish, due to variation in morphology and color in M. kotokui. However, the unraised rugae on the posterodorsal portion of the mesonotum are usually characteristic, though some M. kotokui workers have this character. It is best in the field to examine the body coloration of a number of workers from each colony. Also, M. rubra has fewer strong rugae, and is more smooth on the lower half of the mesonotal pleura than M. kotokui; its propodeal spines are shorter, and the rugae on the anterior basal portion of the 1st gastral tergite are relatively weak and fewer in number. Although Japanese specimens differ morphologically from European M. rubra material, and resemble M. kotokui in petiolar morphology, the name Myrmica rubra is applied to them for the present.
Keys including this Species
- Key to Myrmica of West Europe and North Africa
- Key to Myrmica of East Europe, West Siberia, northern Kazakhstan, Caucasus, Asia Minor, Turkmenistan and Iran
- Key to Myrmica of species of East Siberia, Russian Far East, Mongolia, Korean Peninsula, northern China, and Japan
- Key to Myrmica of the Middle Asian Mountains
- Key to Myrmica of Romania
- Key to Myrmica of China
- Key to Myrmica of North America
- Key to New England Myrmica
- Key to Myrmica of European ambers
Distribution
Europe (in the south mostly in mountains), Siberia, to the east until Transbaikalia, to the north until Forest-Tundra Zone; in Transcaucasus and Middle Asian mountains is quite rare; introduced in northeastern and northwestern United States and western Canada.
In Greece rare species recorded only from northern parts of Macedonia province. New to Thrace (Borowiec et al., 2022).
Latitudinal Distribution Pattern
Latitudinal Range: 70.377854° to 38.983611°.
| North Temperate |
North Subtropical |
Tropical | South Subtropical |
South Temperate |
- Source: AntMaps
Distribution based on Regional Taxon Lists
Nearctic Region: Canada, United States.
Palaearctic Region: Andorra, Armenia, Austria, Azerbaijan, Belarus, Belgium, Bulgaria, China, Croatia, Czechia, Denmark, Estonia, Finland, France, Georgia, Germany, Greece, Hungary, Iberian Peninsula, Japan, Jersey, Kyrgyzstan, Latvia, Liechtenstein, Lithuania, Luxembourg, Mongolia, Montenegro, Netherlands, Norway, Poland, Portugal, North Macedonia, Republic of Moldova, Romania, Russian Federation, Slovakia, Slovenia, Spain, Sweden, Switzerland, Türkiye, Ukraine, United Kingdom of Great Britain and Northern Ireland.
Distribution based on AntMaps
Distribution based on AntWeb specimens
Check data from AntWeb
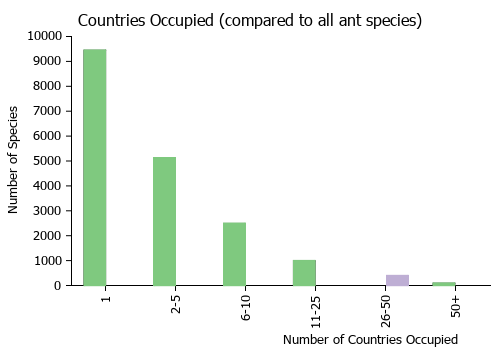
Countries Occupied
| Number of countries occupied by this species based on AntWiki Regional Taxon Lists. In general, fewer countries occupied indicates a narrower range, while more countries indicates a more widespread species. |
|
Biology
Radchenko and Elmes (2010):

M. rubra is a eurytopic species distributed widely throughout Europe and West Siberia where it can dominate some habitats. It thrives in damp habitats, especially soils with high water tables or habitats in areas of high rainfall. However, it is seldom found living in tussocks on true bogs, in the manner of some populations of M. scabrinodis and M. ruginodis. In western Europe it is considered to be a species of damp meadows and is rarely found in woods and forests, the largest populations usually occur on west-facing slopes with heavy clayey (often limestone) soils, where it builds nests in the soil and under flat stones. These sites often have high rainfall and the moisture is held in the heavy soils. In eastern Europe (Russia, Ukraine etc.) it is considered to be more of a forest species inhabiting many different kinds of forests (except those with light, dry soils), where it builds nests in the soil under moss and in or under rotten wood. In central Europe M. rubra is often very abundant in grass on forest, woodland and hedgerow edges, and in Germany, Poland and France etc. it is particularly abundant in the longer vegetation at the edges of water meadows used for haymaking and grazing. Throughout its entire range it is associated with meadows bordering rivers and lakes. In recent years it has become important in nature conservation as the primary ant host of the endangered butterfly species Phengaris nausithous (Bergstrasser) (see papers in Settele et a!. 2005).
M. rubra colonies do not need a long season of high temperatures to complete their life cycles, in most habitats the ants do not become active until the end of April and are entering a pre-hibernation state by late September (Elmes 1982). They have an active basal physiology (compared to many other Myrmica species) that has adapted to local environments in different parts of its range. Habitat selection seems to be determined by a trade-off between sufficient insolation to complete their life cycle and maintaining a high humidity within the soil nest (assuming other factors such as food availability and nest site suitability being equal). Thus at sea level in the more oceanic climates of western Europe, woodlands are too cold in summer while east-facing meadows get too hot and dry whereas in the much more continental climates of eastern Europe, the hot summers enable them to live in woodlands which dessicate less rapidly than open meadows. Mountains of course, make their own local climates so that for example, in the Carpathians populations favour more open meadow habitats at higher altitudes that are ecologically very similar to the prime habitat in western Europe.
Generally, the microhabitat favoured by M. rubra colonies living by rivers and wet meadows ranges from open grass (about 10-20 cm tall) in the north and west of Europe to much longer grass and reeds (1-2 m tall) in southern and eastern habitats. Much less is known about its distribution in West Siberia: M. rubra is one of the commonest ants in various habitats of West and East Siberia (Reznikova 1983; Dmitrienko, Petrenko 1976) and is particularly common in rivers meadows in north-eastern Kazakhstan (M. Woyciechowski, pers. comm.). The principal competitors of M. rubra are other Myrmica species but in meadows it faces strong competition from Lasius niger (Czechowski 1985).
Morphologically M. rubra is comparatively stable over its very wide range (unlike, for example, M. scabrinodis, which is probably undergoing current speciation in Europe – see notes to that species), most of its local adaptation appears to have been physiological and perhaps behavioural. Consequently, given its abundance, it would be an ideal candidate to attempt phylogeographical history of its invasion of Europe using modern molecular analytical techniques. Until this occurs we can only hypothesise from whence M. rubra spread. It seems probable to us, that it survived the last ice age in Middle Asia or maybe the Balkans or southeast Europe, on so-called “tundra-steppes”, and spread rapidly into Europe along the great river margins as the ice melted. Coinciding with man's deforestation of Europe many new habitats were created in the oceanic part of Europe, the forests of which would have been generally too cold for colonisation. Thus in a sense M. rubra is pre-adapted to invade anthropogenic habitats (gardens, agrocoenoses) especially in areas of high rainfall. This might help explain why it has been a very successful invader of the eastern seaboard of USA and Canada.

It is most useful to think of M. rubra in terms of “nests”, colonies can consist of single nests, or polycalic aggregations of several nests and even huge aggregations that can be considered as super-colonies (see Garnas et al. 2007). At a typical meadow site in Southern England nests were on average 1.8 m apart (Elmes 197 4a). Most studies to date show that individual nests contain from a few tens of workers to up to eight thousand with a mean of about 1,000 (Wardlaw and Elmes 1996); nests are usually polygynous with a mean of about 15 queens and nests hardly ever exceed a maximum density of 1 queen per 10 workers (Elmes 1973a). Remarkably average nest size does not differ notably from these estimates even in the super colonies of the USA (Garnas et al. 2007). In wild populations workers “turn-over” quite rapidly and live on average less than 2 years (Brian 1972) and most queens are equally short-lived (Elmes 1980; Seppa and Walin, 1996) .The density of queens in nests varies between populations in space and time (Elmes and Petal 1990), a statistical analysis of weather variables indicated that the nest population of both workers and queens increased as the mean late summer temperatures deviated from about 16°C, which was more or less the long-term average for the sites studied. It was suggested that the most probable explanation was that the local population structure was adapted to the long term conditions at a particular site and in years when conditions deviated from this, nests were less likely to fragment (see below) and so on average, were somewhat larger. This fits with the larger picture for local physiological adaptation by populations of Myrmica in different parts of their range (Elmes et. al. 1999) and the observations in particular for M. rubra (e.g. Brian 1973a; Kipyatkov 1979; Elmes and Wardlaw 1983a; Raybould and Pearson 1992). Nests are usually built into the soil and the design of the excavated nest is tailored to the conditions of the nest site (e.g. Evesham 1992). In coastal meadows M. rubra are less tolerant of salted soils compared to M. scabrinodis but nests can survive periodic inundation (Boomsma and De Vries 1980; Boomsma and lsaaks 1982). Young mated queens are available for much of the active year and are often recruited into nests (Elmes 1982) so that local populations can vary considerable for individual relatedness and level of effective polygyny (Seppa and Walin 1996). Consequently nest fission is a common way of reproduction in M. rubra. Nests frequently fragment in spring and in some circumstances become aggressive towards each other (Czechowski 1984). Often fragments recombine in autumn or they may remain associated as a polycalic colony. In some circumstances M. rubra populations may form huge polycalic colonies with hundreds of thousands or even millions of workers, like in some North America populations and very occasionally in some European populations (personal observation). In such populations it appears that there is a high level of food availability, nests grow rapidly and fragment but do not need to move far from the parent nest. This results in a very dense population of nests though individual nest sizes remain on average about 1000 workers. The proclivity for fragmenting into small colonies each containing several queens seems to be a useful adaptation to rapidly invading new habitat and at the same time incipient colonies are easily spread and introduced into new habitats by man's activities (in plant pots, roots of transplanted shrubs and trees – see Groden et al. 2005 and Garnas et. al. 2007).

Although workers in supercolonies appear to be very tolerant towards their neighbouring colony members, they can be very aggressive towards other organisms, stinging people and other animals freely (personal observations); however, laboratory studies suggest that M. rubra workers do not have especially different aggressive responses compared to other members of the genus (De Vroey and Pasteels 1978). M. rubra have a well equipped sting apparatus (Billen 1986) and although some people react allergically to the venom, the sting and venom does not seem markedly different from that of other free-stinging myrmicines (e.g. Blum and Herman 1978). Most people (including one of us - AR) think that M. rubra stings seem particularly painful compared to other Myrmica species. However in the opinion of the other author (GWE) when individuals of other Myrmica are provoked into stinging (usually in hot conditions) a single sting can be equally painfully as that of M. rubra, perhaps even more so if the specimen is large: M. rubra having acquired its reputation because colonies are corporately aggressive and individuals sting rapidly and frequently even when fairly cool.
M. rubra is a generalist scavenger and predator hunting various small invertebrates (e.g. Petal 1967), but also utilize honeydew and nectar (flowers and extrafloral nectaries, e.g. Felton 1959), aphids and scale insects. They forage on trees and shrubs more frequently than any other Myrmica species (except perhaps M. ruginodis); though in Europe arboreal foraging is quite rare while in the supercolonies of Maine, USA very many M. rubra workers can be seen foraging high into the canopy (personal observations). M. rubra workers often forage in groups (e.g. Dlussky et al. 1978) and they lay and follow chemical foraging trails (e.g. Cammaerts-Tricot and Verhaeghe 1974; Cammaerts-Tricot et al. 1977). Single foragers (weighing about 2 mg) are able to exert pulling-forces of about 100 mg. developing a mean power of about 5.8 ergs/s (Sudd 1965), a third to a quarter of the strength and power exerted by Formica lugubris Zett. workers.
Winged sexuals (gynes and males) are produced in June and “mature” inside the nest until July. The ontogeny of larval development and caste determination has been extensively studied by M. V. Brian using M. rubra as a model species and has been found to be very complex; for example, the hormonal state of the queen influences larval hormone production and ontogeny (Brian 1959, 1974), trophic conditions are involved (Brian and Abbott 1977) as are the age and numbers of workers (Brian and Jones 1980), seasonality has an effect (Pearson and Raybould 1997) and even gut parasite load might have some impact (Pearson and Raybould 1998). A model based on these interactions show that gyne production might be periodic (Brian et al. 1981) and this may account for the observation that the mean size of workers and queens in nests is positively correlated with worker number and negatively correlated with queen number (Elmes 1974b).
Nuptial flights occur from late July and have been reported as late as October. Compared to many other Myrmica species, M. rubra mating swarms can be quite large aggregations and they have frequently been reported flying quite long distances to join swarms on church towers, high trees and mountain-tops (e.g. Hubbard and Nagell 1976; Woyciechowski 1990b; personal observations). We have on occasion observed nests having recruited a mixture of their own daughters and other young queens (all fertilised), but we are not sure whether their daughters mated in or near to the nest prior to joining the parent colony or flew to a distant swarm, mated and found their way home again. While the latter seems improbable it is what happens in the case of honey bees.
Bertelsmeier et al. (2015) examined elements of interspecific aggression between this species and several other highly invasive ants. In laboratory assays Myrmica rubra was adept at avoiding aggressive interactions. When confronted by workers of other invasive ant species M. rubra either acted indifferently or moved away.
Foraging/Diet
See the general biology discussion above for an overview of diet and foraging. Novgorodova (2015b) investigated ant-aphid interactions of a dozen honeydew collecting ants in south-central Russia. All of the ants studied had workers that showed high fidelity to attending particular aphid colonies, i.e, individual foragers that collect honeydew tend to return to the same location, and group of aphids, every time they leave the nest. Myrmica rubra showed no specialization beyond this foraging site fidelity. Foragers tended Chaitophorus populeti (Panzer) and Aphis pomi De Geer.
Bologna and Detrain (2015) examined foraging behavior in a laboratory experiment with M. rubra obtained from locations in Belgium. They found that the ants became satiated and showed a large decline over time in retrieval of elaiosome bearing seeds of Viola odorata. Seeds were offered once a week for 5 consecutive weeks and again at week 12. A similar experiment with dead fruit flies showed a consistent foraging response where the ants collected most of the offered fruit flies.
Guiliani et al. (2019) observed this species foraging on extrafloral nectaries of the invasive Reynoutria x bohemica (Polygonaceae) in Tuscany. The habitats examined were river banks and disturbed habitats.
Introduced Range Studies
British Columbia, Canada
Naumann and Higgins (2015) examined the influence of this species on native insects. Abstract: Pitfall trapping revealed that the European fire ant, Myrmica rubra (Linnaeus) (Hymenoptera: Formicidae), represents an unusual example of a temperate invasive ant species. In British Columbia, Canada, M. rubra populations are associated with a decreased incidence and abundance of other ant species in three different plant communities when compared with M. rubra-free control areas. M. rubra represented more than 99.99% of the total ant fauna caught in the infested areas, and the numbers of M. rubra captured in the plant communities ranged from over 10 times to over 1300 times the total number of all ants collected in corresponding M. rubra-free areas. Total numbers of some taxa of insects and non-insect arthropods, including those likely to be competitors or prey of M. rubra, were reduced where the invasive species was present. Biodiversity indexes for the overall suite of captured arthropod species were lower where M. rubra was present in all three plant communities but most of this decrease can be attributed to the difference in the ant fauna.
Naumann et al. (2017) investigated molecular genetic diversity and found there appear to be just a few supercolonies and the level of aggression between individuals across separate locations is very low.
Experimental Studies - Social Immunity
Diez et al. (2015), a study examining pathogens and colony hygiene - Abstract: Ants have developed prophylactic and hygienic behaviours in order to limit risks of pathogenic outbreaks inside their nest, which are often called social immunity. Here, we test whether ants can adapt the “social immune response” to the level of pathogenic risk in the colony. We challenged Myrmica rubra colonies with dead nestmates that had either died from being frozen or from infection by the fungus Metarhizium anisopliae. Ant survival was compromised by the presence of the fungus-bearing corpses: workers died faster with a significantly lower survival from the 4th day compared to workers challenged with freeze-killed corpses. When faced with fungus-bearing corpses, workers responded quickly by increasing hygienic behaviours: they spent more time cleaning the nest, moving the corpses, and self-grooming. Ants in fungus-threatened colonies also decreased contact rates with other workers, and moved corpses further in the corners of the nest than in colonies in contact with non-infected corpses. These results show that ant colonies are able to assess the risk level associated with the presence of corpses in the nest, and adjust their investment in terms of hygienic behaviour.
Association with Other Organisms
 Explore: Show all Associate data or Search these data. See also a list of all data tables or learn how data is managed.
Explore: Show all Associate data or Search these data. See also a list of all data tables or learn how data is managed.
- This species is a xenobiont for the ant Formica fusca (a xenobiont) in Finland, United Kingdom (Czechowski, W. 2003; Czechowski, 2004; Kanizsai et al., 2013; Morley, 1945) (habitat foreshore, nests in decaying wood, under stone).
- This species is a xenobiont for the ant Lasius niger (a xenobiont) in Finland (Czechowski, 2004; Kanizsai et al., 2013) (Shore meadow; at road. Under stone; between asphalt edge and grass.).
- This species is a xenobiont for the ant Lasius platythorax (a xenobiont) in Finland (Czechowski, 2004; Kanizsai et al., 2013) (Forest. In decaying wood).
- This species is a xenobiont for the ant Leptothorax muscorum (a xenobiont) in Finland (Czechowski, 2004; Kanizsai et al., 2013) (Shore meadow. Under stone).
Hymenoptera
- This species is a host for the pteromalid wasp Asaphes vulgaris (a parasite) (Universal Chalcidoidea Database) (associate).
- This species is a host for the pteromalid wasp Pachyneuron aphidis (a parasite) (Universal Chalcidoidea Database) (associate).
- This species is a associate (details unknown) for the encyrtid wasp Syrphophagus aphidivorus (a associate (details unknown)) (Quevillon, 2018).
- This species is a host for the encyrtid wasp Syrphophagus aphidivorus (a parasite) (Universal Chalcidoidea Database) (associate).
Hemiptera
- This species is a mutualist for the aphid Cinara apini (a trophobiont) (Jones, 1927; Saddiqui et al., 2019).
Mites
- This species is a host for the mite Petalomium podolicus (a parasite) (Khaustov, 2015) (ectoparasite).
- This species is a host for the mite Scutacarus longisetus (a parasite) (Khaustov, 2015) (ectoparasite; as Myrmica laevinodis).
Nematode
- This species is a host for the nematode Mermithidae (unspec.) (a parasite) in Poland (Czechowsk et al., 2007; Laciny, 2021).
- This species is a host for the nematode Mermithidae (unspecified "Mermix") (a parasite) in Germany (Gosswald, 1930; Laciny, 2021).
Cestoda
- This species is a host for the cestode Davaineidae (Raillietina urogalli??) (a parasite) in Scotland (Muir, 1954; Laciny, 2021).
- This species is a host for the cestode Raillietina urogalli (a parasitoid) (Quevillon, 2018) (encounter mode secondary; indirect transmission; transmission outside nest).
Gregarine
- This species is a host for the gregarine Mattesia geminata (a parasite) in Germany (Buschinger & Kleespies, 1999).
Fungi
- This taxon is a host for the fungi Ophiocordyceps myrmecophila (Shrestha et al., 2017) and Rickia wasmanniii (Espadaler & Santamaria, 2012).
- This species is a host for the fungus Ophiocordyceps kniphofioides (a parasitoid) (Quevillon, 2018) (encounter mode primary; direct transmission; transmission outside nest).
- This species is a host for the fungus Ophiocordyceps myrmicarum (a parasitoid) (Quevillon, 2018) (encounter mode primary; direct transmission; transmission outside nest).
- This species is a host for the fungus Ophiocordyceps subramanianii (a parasitoid) (Quevillon, 2018) (encounter mode primary; direct transmission; transmission outside nest).
- This species is a host for the fungus Rickia wasmannii (a parasite) (Quevillon, 2018) (encounter mode primary; direct transmission; transmission within nest).
- This species is a host for the fungus Rickia wasmannii (a parasite) (Quevillon, 2018) (encounter mode primary; direct transmission; transmission within nest).
Flight Period
| X | |||||||||||
| Jan | Feb | Mar | Apr | May | Jun | Jul | Aug | Sep | Oct | Nov | Dec |
Source: antkeeping.info.
- Check details at Worldwide Ant Nuptial Flights Data, AntNupTracker and AntKeeping.
- Explore: Show all Flight Month data or Search these data. See also a list of all data tables or learn how data is managed.
Life History Traits
- Queen number: polygynous (Rissing and Pollock, 1988; Frumhoff & Ward, 1992)
- Queen type: winged (Rissing and Pollock, 1988; Frumhoff & Ward, 1992) (queen-right worker reproduction)
- Queen mating frequency: multiple (Rissing and Pollock, 1988; Frumhoff & Ward, 1992)
- Worker-produced males: present (Smeeton, 1981; Frumhoff & Ward, 1992)
- Mean colony size: 1,000 (Stradling, 1970; Petal, 1972; Cammaerts & Cammaerts, 1980; Beckers et al., 1989)
- Foraging behaviour: mass recruiter (Stradling, 1970; Petal, 1972; Cammaerts & Cammaerts, 1980; Beckers et al., 1989)
Castes
Worker
Images from AntWeb
   
| |
| Worker. Specimen code casent0010684. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by UCDC, Davis, CA, USA. |
   
| |
| Worker. Specimen code casent0104804. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by UCDC, Davis, CA, USA. |
   
| |
| Worker. Specimen code casent0172750. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by CAS, San Francisco, CA, USA. |
   
| |
| Worker. Specimen code casent0010644. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by CAS, San Francisco, CA, USA. |
Queen
Images from AntWeb
   
| |
| Queen (alate/dealate). Specimen code casent0172707. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by CAS, San Francisco, CA, USA. |
    
| |
| Queen (alate/dealate). Specimen code casent0173172. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by CAS, San Francisco, CA, USA. |
Male
Images from AntWeb
     
| |
| Male (alate). Specimen code casent0172751. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by CAS, San Francisco, CA, USA. |
     
| |
| Queen (alate/dealate). Specimen code casent0173173. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by CAS, San Francisco, CA, USA. |
    
| |
| Male (alate). Specimen code casent0104805. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by UCDC, Davis, CA, USA. |
Nomenclature
The following information is derived from Barry Bolton's Online Catalogue of the Ants of the World.
- rubra. Formica rubra Linnaeus, 1758: 580 (w.) EUROPE. Latreille, 1802c: 248 (q.m.); Wheeler, G.C. & Wheeler, J. 1953a: 118 (l.); Hauschteck, 1965: 325 (k.). Combination in Myrmica: Latreille, 1804: 179. Senior synonym of laevinodis (and its junior synonyms bruesi, champlaini, longiscapus): Yarrow, 1955b: 114; Arnol'di, 1970b: 1839; Boven, 1977: 115; Arnol'di & Dlussky, 1978: 530; Collingwood, 1979: 52; Seifert, 1988b: 5; of europaea: Radchenko, Czechowski & Czechowska, 1997: 483; Czechowski, Radchenko & Czechowska, 2002: 17; of microrubra: Steiner, Schlick-Steiner, Konrad, et al. 2006: 777. Current subspecies: nominal plus khamensis, neolaevinodis. See also: Emery, 1908a: 169; Brian, M.V. & Brian, A.D. 1949: 393; Smith, D.R. 1979: 1350; Pearson, 1981: 75; Seifert, 1988b: 5; Onoyama, 1989a: 131; Atanassov & Dlussky, 1992: 83; Radchenko & Elmes, 2010: 228.
- laevinodis. Myrmica laevinodis Nylander, 1846a: 927, pl. 18, figs. 5, 31 (w.q.m.) FINLAND. Subspecies of rubra: Forel, 1874: 76; Emery & Forel, 1879: 460; Wheeler, W.M. 1906d: 315; Emery, 1914d: 156; Forel, 1915d: 28; Menozzi, 1918: 82; Karavaiev, 1927c: 259; Creighton, 1950a: 104. Status as species: Saunders, E. 1880: 215; André, 1883a: 316; Dalla Torre, 1893: 110; Ruzsky, 1905b: 662; Bondroit, 1912: 351; Donisthorpe, 1915d: 110; Bondroit, 1918: 104; Müller, 1923: 40; Finzi, 1926: 83; Stitz, 1939: 78; Holgersen, 1940: 184; Novak & Sadil, 1941: 76; Weber, 1947: 441; Bernard, 1967: 119; Baroni Urbani, 1971c: 22; Kutter, 1977c: 65. Senior synonym of longiscapus: Mayr, 1863: 433; of champlaini: Smith, M.R. 1951a: 789; of bruesi: Creighton, 1950a: 104. Junior synonym of rubra: Yarrow, 1955b: 114; Arnol'di, 1970b: 1839; Arnol'di & Dlussky, 1978: 530; Collingwood, 1979: 52; Seifert, 1988b: 5; Radchenko, 2007: 30.
- europaea. Myrmica laevinodis var. europaea Finzi, 1926: 84 (w.) NORWAY. [First available use of Myrmica rubra subsp. champlaini var. europaea Forel, 1911h: 457; unavailable name.] Santschi, 1931b: 339 (m.). Subspecies of laevinodis: Stitz, 1939: 83; of rubra: Bolton, 1995b: 279. Junior synonym of rubra: Radchenko, Czechowski & Czechowska, 1997: 483; Czechowski, Radchenko & Czechowska, 2002: 17.
- bruesi. Myrmica laevinodis var. bruesi Weber, 1947: 453 (w.q.m.) U.S.A. [First available use of Myrmica rubra subsp. laevinodis var. bruesi Wheeler, W.M. 1906a: 38; unavailable name.] Junior synonym of laevinodis: Creighton, 1950a: 104.
- champlaini. Myrmica rubra r. champlaini Forel, 1901h: 80 (w.) CANADA. Subspecies of laevinodis: Weber, 1947: 454; of rubra: Creighton, 1950a: 103. Junior synonym of laevinodis: Smith, M.R. 1951a: 789.
- longiscapus. Myrmica longiscapus Curtis, 1854: 213, pl. 23, figs. 11-14 (w.q.m.) GREAT BRITAIN. Junior synonym of laevinodis: Mayr, 1863: 433.
- microrubra. Myrmica microrubra Seifert, 1993: 10, figs. 1, 4 (q.m.) GERMANY. Junior synonym of rubra: Steiner, Schlick-Steiner, Konrad, et al. 2006: 777. See also: Czechowski, Radchenko & Czechowska, 2002: 19; Radchenko & Elmes, 2003a: 236.
Unless otherwise noted the text for the remainder of this section is reported from the publication that includes the original description.
Description

{kind=link}
Karyotype
- See additional details at the Ant Chromosome Database.
- Explore: Show all Karyotype data or Search these data. See also a list of all data tables or learn how data is managed.
- 2n = 48 (Switzerland) (Hauschteck-Jungen & Jungen, 1983) (as Myrmica laevinodis).
- n = 23, 2n = 46 (Japan) (Imai, 1969).
Etymology
Radchenko and Elmes (2010) - from the Latin word rubra = red, to describe its generally reddish colour.
References
- Adams, R.M.M., Wells, R.L., Yanoviak, S.P., Frost, C.J., Fox, E.G.P. 2020. Interspecific Eavesdropping on Ant Chemical Communication. Frontiers in Ecology and Evolution 8. (doi:10.3389/fevo.2020.00024).
- Alvarez-Blanco, P., Broggi, J., Cerdá, X., González-Jarri, O., Angulo, E. 2020. Breeding consequences for a songbird nesting in Argentine ant’ invaded land. Biological Invasions 22, 2883–2898. (doi:10.1007/S10530-020-02297-3).
- Aranda-Rickert, A., Fracchia, S. 2012. La biología de Pogonomyrmex cunicularius pencosensis (Hymenoptera: Formicidae) en relación con su comportamiento como dispersora de semillas con eleosoma en el Noroeste semiárido argentino. Revista de la Sociedad Entomológica Argentina 71 (1-2): 11-27.
- Arnol'di, K. V. 1970b. Review of the ant genus Myrmica (Hymenoptera, Formicidae) in the European part of the USSR. Zool. Zh. 4 49: 1829-1844 (page 1839, Senior synonym of laevinodis (and its junior synonyms bruesi, champlaini and longiscapus))
- Arnol'di, K. V.; Dlussky, G. M. 1978. Superfam. Formicoidea. 1. Fam. Formicidae - ants. Pp. 519-556 in: Medvedev, G. S. (ed.) Keys to the insects of the European part of the USSR. Vol. 3. Hymenoptera. Part 1. Opredeliteli Faune SSSR 119:3-584. (page 530, Senior synonym of laevinodis (and its junior synonyms bruesi, champlaini and longiscapus))
- Atanassov, N.; Dlussky, G. M. 1992. Fauna of Bulgaria. Hymenoptera, Formicidae. Fauna Bûlg. 22: 1-310 (page 83, see also)
- Ataya, H., Lenoir, A. 1984. Le comportement necrophorique chez la formi Lasius niger L. Insectes Sociaux 31: 20-33.
- Baer, B. 2011. The copulation biology of ants (Hymenoptera: Formicidae). Myrmecological News 14: 55-68.
- Barbero, F., Patricelli, D., Witek, M., Balletto, E., Casacci, L.P., Sala, M., Bonelli, S. 2012. Myrmica Ants and Their Butterfly Parasites with Special Focus on the Acoustic Communication. Psyche: A Journal of Entomology 2012, 1–11 (doi:10.1155/2012/725237).
- Báthori, F., Rádai, Z., Tartally, A. 2017. The effect of Rickia wasmanniii (Ascomycota, Laboulbeniales) on the aggression and boldness of Myrmica scabrinodis (Hymenoptera, Formicidae). Journal of Hymenoptera Research. 58:41–52. (doi:10.3897/jhr.58.13253)
- Baty, J.W., Bulgarella, M., Dobelmann, J., Felden, A., Lester, P.J. 2020. Viruses and their effects in ants (Hymenoptera: Formicidae). Myrmecological News 30: 213-228 (doi:10.25849/MYRMECOL.NEWS_030:213).
- Beckers R., Goss, S., Deneubourg, J.L., Pasteels, J.M. 1989. Colony size, communication and ant foraging Strategy. Psyche 96: 239-256 (doi:10.1155/1989/94279).
- Bernadou, A., Fourcassié, V., Espadaler, X. 2013. A preliminary checklist of the ants (Hymenoptera, Formicidae) of Andorra. ZooKeys 277, 13–23 (doi:10.3897/zookeys.277.4684).
- Bertelsmeier, C., A. Avril, O. Blight, A. Confais, L. Diez, H. Jourdan, J. Orivel, N. St Germes, and F. Courchamp. 2015. Different behavioural strategies among seven highly invasive ant species. Biological Invasions. 17:2491-2503. doi:10.1007/s10530-015-0892-5
- Berton, F., Lenoir, A., Newbahari, E., Barreau, S. 1991. Ontogeny of queen attraction to workers in the ant Cataglyphis cursor (Hymenoptera: Formicidae). Insectes Sociaux 38: 293-305.
- Bezděčková, K., Bezděčka, P., Macek, J., Malenovský, I. 2017. Catalogue of type specimens of ants (Hymenoptera: Formicidae) deposited in Czech museums. Acta Entomologica Musei Nationalis Pragae 57(1): 295-308 (doi:10.1515/aemnp-2017-0076).
- Bologna, A. and C. Detrain. 2015. Steep Decline and Cessation in Seed Dispersal by Myrmica rubra Ants. Plos One. 10. doi:10.1371/journal.pone.0139365
- Bologna, A., Detrain, C. 2019. Larvae promote a decline in the harvesting of myrmecochorous seeds by Myrmica rubra ants. Insectes Sociaux 66, 453–461 (doi:10.1007/s00040-019-00706-y).
- Boomsma, J.J., Leusink, A. 1981. Weather conditions during nuptial flights of four European ant species. Oecologia 50, 236–241. (doi:10.1007/bf00348045).
- Borowiec, L. 2014. Catalogue of ants of Europe, the Mediterranean Basin and adjacent regions (Hymenoptera: Formicidae). Genus (Wroclaw) 25(1-2): 1-340.
- Borowiec, L., Lebas, C., Salata, S. 2022. Notes on ants (Hymenoptera: Formicidae) from three northern Aegean islands – Lemnos, Samothraki and Thasos. Annals of the Upper Silesian Museum in Bytom, Entomology 31: 1-14 (doi:10.5281/ZENODO.7346453).
- Borowiec, L., Salata, S. 2022. A monographic review of ants of Greece (Hymenoptera: Formicidae). Vol. 1. Introduction and review of all subfamilies except the subfamily Myrmicinae. Part 1: text. Natural History Monographs of the Upper Silesian Museum 1: 1-297.
- Borowiec, M.L., Cover, S.P., Rabeling, C. 2021. The evolution of social parasitism in Formica ants revealed by a global phylogeny. Proceedings of the National Academy of Sciences 118, e2026029118 (doi:10.1073/pnas.2026029118).
- Boulay, R., Cerda, X., Fertin, A., Ichinose, K., Lenior, A. 2009. Brood development into sexual females depends on the presence of a queen but not on temperature in an ant dispersing by colony fission, Aphaenogaster senilis. Ecological Entomology 34, 595–602 (doi:10.1111/j.1365-2311.2009.01108.x).
- Boulay, R., Quagebeur, M., Godzinska, E.J., Lenoir, A. 1999. Social isolation in ants: Evidence of its impact on survivorship and behavior in Camponotus fellah (Hymenoptera, Formicidae).
- Boven, J. K. A. van. 1977. De mierenfauna van België (Hymenoptera: Formicidae). Acta Zool. Pathol. Antverp. 67: 1-191 (page 115, Senior synonym of laevinodis (and its junior synonyms bruesi, champlaini and longiscapus))
- Bracko, G., Wagner, H.C., Schulz, A., Gioahin, E., Maticic, J., Trantnik, A. 2014. New investigation and a revised checklist of the ants (Hymenoptera: Formicidae) of the Republic of Macedonia. North-Western Journal of Zoology 10: 10-24.
- Brian, M. V.; Brian, A. D. 1949. Observations on the taxonomy of the ants Myrmica rubra L. and M. laevinodis Nylander. (Hymenoptera: Formicidae.). Trans. R. Entomol. Soc. Lond. 100: 393-409. (page 393, see also)
- Brian, M.V. 1956. Group form and causes of working inefficiency in the ant Myrmica rubra. Physiological Zoology 29: 173-194.
- Brown, M.J.F., Bonhoeffer, S. 2003. On the evolution of claustral colony founding in ants. Evolutionary Ecology Research 5: 305–313.
- Buschinger, A., Kleespies, R.G. 1999. Host range and host specificity of an ant-pathogenic gregarine parasite, Mattesia geminata (Neogregarinida: Lipotrophidae). Entomologia Generalis 24: 93-104 (doi:10.1127/entom.gen/24/1999/93).
- Cantone S. 2017. Winged Ants, The Male, Dichotomous key to genera of winged male ants in the World, Behavioral ecology of mating flight (self-published).
- Carroll, T.M. 2011. The ants of Indiana (Hymenoptera: Formicidae). M.S. thesis, Purdue University.
- Castracani, C., Spotti, F.A., Schifani, E., Giannetti, D., Ghizzoni, M., Grasso, D.A., Mori, A. 2020. Public engagement provides first insights on Po Plain ant communities and reveals the ubiquity of the cryptic species Tetramorium immigrans (Hymenoptera, Formicidae). Insects 11, 678. (doi:10.3390/insects11100678).
- Cerda, X., Arnan, X., Retana, J. 2013. Is competition a significant hallmark of ant (Hymenoptera: Formicidae) ecology? Myrmecological News 18: 131-147.
- Choppin, M., Graf, S., Feldmeyer, B., Libbrecht, R., Menzel, F., Foitzik, S. 2021. Queen and worker phenotypic traits are associated with colony composition and environment in Temnothorax rugatulus (Hymenoptera: Formicidae), an ant with alternative reproductive strategies. Myrmecological News 31: 61-69 (doi:10.25849/MYRMECOL.NEWS_031:061).
- Chung, F.-Y., Hsu, P.-W., Hsu, F.-C., Lin, C.-C. 2025. A new genus in a hyperdiverse ant lineage (Hymenoptera: Formicidae: Myrmicinae) revealed by integration of morphological and molecular evidence. Myrmecological News 35: 29-60 (doi:10.25849/MYRMECOL.NEWS_035:029).
- Collingwood, C. A. 1979. The Formicidae (Hymenoptera) of Fennoscandia and Denmark. Fauna Entomol. Scand. 8: 1-174 (page 52, Senior synonym of laevinodis (and its junior synonyms bruesi, champlaini and longiscapus))
- Collingwood, C.A., Prince, A. 1998. A guide to ants of Continental Portugal (Hymenoptera: Formicidae). Boletim da Sociedade Portuguesa de Entomologia. Supl nº5, pp 49.
- Csata, E., Czekes, Z., Eros, K., Nemet, E., Hughes, M., Csosz, S., Marko, B. 2013. Comprehensive survey of Romanian myrmecoparasitic fungi: new species, biology and distribution. North-western Journal of Zoology 9: 23-29.
- Csősz, S., Báthori, F., Gallé, L., Lőrinczi, G., Maák, I., Tartally, A., Kovács, É., Somogyi, A.Á., Markó, B. 2021. The myrmecofauna (Hymenoptera: Formicidae) of Hungary: Survey of ant species with an annotated synonymic inventory. Insects 16;12(1):78 (doi:10.3390/insects12010078).
- Csosz, S., Marko, B., Galle, L. 2011. The myrmecofauna (Hymenoptera: Formicidae) of Hungary: an updated checklist. North-Western Journal of Zoology 7: 55-62.
- Cuvillier-Hot, V., Salin, K., Devers, S., Tasiemski, A., Schaffner, P., Boulay, R., Billiard, S., Lenoir, A. 2014. Impact of ecological doses of the most widespread phthalate on a terrestrial species, the ant Lasius niger. Environmental Research 131, 104–110 (doi:10.1016/j.envres.2014.03.016).
- Czechowski, W., Czechowska, W., Radchenko, A. 2007. Strikingly malformed host morphology: Myrmica rugulosa Nyl. and Myrmica sabuleti Mein. (Hymenoptera: Formicidae) parasitised by mermithid nematodes. Fragmenta Faunistica 50: 139–148.
- Czechowski, W., Radchenko, A., Czechowska, W. 2002. The ants (Hymenoptera, Formicidae) of Poland. MIZ PAS Warsaw.
- Czechowski, W., Rutkowski, T., Stephen, W., Vepsäläinen, K. 2016. Living beyond the limits of survival: wood ants trapped in a gigantic pitfall. Journal of Hymenoptera Research 51, 227–239 (doi:10.3897/jhr.51.9096).
- Czechowski, W.; Radchenko, A.; Czechowska, W. 2002. The ants (Hymenoptera, Formicidae) of Poland. Warsaw: Museum and Institute of Zoology, Polish Academy of Sciences: 200 pp. (page 17, senior synonym of europea)
- Czekes, Z., Radchenko, A.G., Csosz, S., sz-Len, A.S., Tausan, I., Benedek, K., Mark, B. 2012. The ant genus Myrmica Latreille, 1804 (Hymenoptera: Formicidae) in Romania: distribution of species and key for their identification. Entomologica Romanica 17: 29-50.
- Dahbi, A., Retana, J., Lenoir, A., Cerdá, X. 2007. Nest-moving by the polydomous ant Cataglyphis iberica. Journal of Ethology 26, 119–126 (doi:10.1007/S10164-007-0041-4).
- de Bekker, C., Will, I., Das, B., Adams, R.M.M. 2018. The ants (Hymenoptera: Formicidae) and their parasites: effects of parasitic manipulations and host responses on ant behavioral ecology. Myrmecological News 28: 1-24 (doi:10.25849/myrmecol.news_028:001).
- de la Mora, A., Sankovitz, M., Purcell, J. 2020. Ants (Hymenoptera: Formicidae) as host and intruder: recent advances and future directions in the study of exploitative strategies. Myrmecological News 30: 53-71 (doi:10.25849/MYRMECOL.NEWS_030:053).
- Degueldre, F., Mardulyn, P., Kuhn, A., Pinel, A., Karaman, C., Lebas, C., Schifani, E., Bračko, G., Wagner, H.C., Kiran, K., Borowiec, L., Passera, L., Abril, S., Espadaler, X., Aron, S. 2021. Evolutionary history of inquiline social parasitism in Plagiolepis ants. Molecular Phylogenetics and Evolution 155, 107016 (doi:10.1016/j.ympev.2020.107016).
- Dekoninck, W., Ignace, D., Vankerkhoven, F., Wegnez, P. 2012. Verspreidingsatlas van de mieren van België. Bulletin de la Société royale belge d’Entomologie 148: 95-186.
- Della Santa, E. 2000. L'identification des espèces du genre Myrmica Latreille (Formicidae) de Suisse; essai de présentation synoptique. Bull. Romand Entomol. 18: 169-187 (page 171, status)
- Diez, L., L. Urbain, P. Lejeune, and C. Detrain. 2015. Emergency measures: Adaptive response to pathogen intrusion in the ant nest. Behavioural Processes. 116:80-86. doi:10.1016/j.beproc.2015.04.016
- Dolezal, A.G. 2010. Caste determination in arthropods, In Encyclopedia of Animal Behavior, edited by Michael D. Breed and Janice Moore, Academic Press, Oxford, pages 247-253.
- Dolezal, A.G. 2019. Caste determination in arthropods. In: Encyclopedia of Animal Behavior, 2nd edition, Volume 4: 691–698 (doi:10.1016/B978-0-12-809633-8.20815-7).
- Dubovikoff, D.A., Yusupov, Z.M. 2017. Family Formicidae - Ants. In Belokobylskij S. A. and A. S. Lelej: Annotated catalogue of the Hymenoptera of Russia. Proceedingss of the Zoological Institute of the Russian Academy of Sciences 6: 197-210.
- Duthie, C., Timms, L., Fraser, G.S. 2021. The European fire ant, Myrmica rubra (Linnaeus) (Hymenoptera: Formicidae), in the Credit River watershed. Journal of the Entomological Society of Ontario 152: 15-28.
- Elmes, G. W. 1973. Miniature queens of the ant Myrmica rubra L. (Hymenoptera, Formicidae). Entomologist 106: 133-136.
- Emery, C. 1908a. Beiträge zur Monographie der Formiciden des paläarktischen Faunengebietes. Dtsch. Entomol. Z. 1908: 165-205 (page 169, see also)
- Emmert, W. 1968. Die Postembryonalentwicklung sekretorischer Kopfdrüsen von Formica pratensis Retz. und Apis mellifica L. (Ins. Hym.). Zeitschrift für Morphologie der Tiere 63:1-62.
- Espadaler, X., Santamaria, S. 2012. Ecto- and Endoparasitic Fungi on Ants from the Holarctic Region. Psyche Article ID 168478, 10 pages (doi:10.1155/2012/168478).
- Fedoseeva, E. B. 2015. A technological approach to the description of group foraging in ant Myrmica rubra. Zoologichesky Zhurnal. 94:1163-1178. doi:10.1134/S0013873815080060
- Fournier, D., de Biseau, J.-C., De Laet, S., Lenoir, A., Passera, L., Aron, S. 2016. Social structure and genetic distance mediate nestmate recognition and aggressiveness in the facultative polygynous ant Pheidole pallidula. PLOS ONE 11, e0156440. (doi:10.1371/journal.pone.0156440).
- Giuliani, C., L. Lastrucci, L. Cresti, G. Santini, B. Fogg, and M. M. Lippi. 2019. The morphology and activity of the extrafloral nectaries in Reynoutria x bohemica (Polygonaceae). Plant Biology. 21:975-985. doi:10.1111/plb.13004
- Glaser, F. 2016. Artenspektrum, Habitatbindung und naturschutzfachliche Bedeutung von Ameisen (Hymenoptera, Formicidae) am Stutzberg (Vorarlberg, Österreich). inatura – Forschung 34: 26 S.
- Gospocic, J., Glastad, K.M., Sheng, L., Shields, E.J., Berger, S.L., Bonasio, R. 2021. Kr-h1 maintains distinct caste-specific neurotranscriptomes in response to socially regulated hormones. Cell 184, 5807–5823.e14 (doi:10.1016/j.cell.2021.10.006).
- Gösswald, K. 1930. Weitere Beiträge zur Verbreitung der Mermithiden bei Ameisen. Zoologischer Anzeiger 90: 13-27.
- Groden, E., Drummond, F.A., Garnas, J., Franceour, A. 2005. Distribution of an Invasive Ant, Myrmica rubra (Hymenoptera: Formicidae), in Maine. Journal of Economic Entomology 98(6), 1774-1784.
- Haelewaters, D., Boer, P., Noordijk, J. 2015. Studies of Laboulbeniales (Fungi, Ascomycota) on Myrmica ants: Rickia wasmanniii in the Netherlands. Journal of Hymenoptera Research 44, 39–47 (doi:10.3897/jhr.44.4951).
- Hauschteck, E. 1965. Halbe haploide Chromosomenzahl im Hoden von Myrmica sulcinodis Nyl. (Formicidae). Experientia (Basel) 21: 323-325 (page 325, Karyotype described)
- Horna-Lowell, E., Neumann, K.M., O’Fallon, S., Rubio, A., Pinter-Wollman, N. 2021. Personality of ant colonies (Hymenoptera: Formicidae) – underlying mechanisms and ecological consequences. Myrmecological News 31: 47-59 (doi:10.25849/MYRMECOL.NEWS_031:047).
- Ichinose, K., Lenoir, A. 2009. Reproductive conflict between laying workers in the ant Aphaenogaster senilis. Journal of Ethology 27, 475–481. (doi:10.1007/s10164-008-0145-5).
- Jaisson, P. 1969. Étude de la distribution des organes sensoriels de l'antenne et de leurs relations possibles avec le comportement chez deux fourmis myrmicines: Myrmica lævinodis Nyl. et Aphænogaster gibbosa Latr. récoltées dans la région des Eyzies. Insectes Sociaux 16, 279–312 (doi:10.1007/bf02309000).
- Jansen, G., Savolainen, R. 2010. Molecular phylogeny of the ant tribe Myrmicini (Hymenoptera: Formicidae). Zoological Journal of the Linnean Society 160(3), 482–495 (doi:10.1111/j.1096-3642.2009.00604.x).
- Jansen, G., Savolainen, R., Vepsäläinen, K. 2010. Phylogeny, divergence-time estimation, biogeography and social parasite–host relationships of the Holarctic ant genus Myrmica (Hymenoptera: Formicidae). Molecular Phylogenetics and Evolution 561, 294–304 (doi:10.1016/j.ympev.2010.01.029).
- Kanizsai, O., Lőrinczi, G., Gallé, L. 2013. Nesting associations without interdependence: A preliminary review on plesiobiosis in ants. Psyche 2013, 238602 (doi:10.1155/2013/238602).
- Kiran, K., Karaman, C. 2020. Additions to the ant fauna of Turkey (Hymenoptera, Formicidae). Zoosystema 42(18), 285-329 (doi:10.5252/zoosystema2020v42a18).
- Kirchmair, G., Friess, T. et al. 2017. Zoologischer Bericht vom Tag der Biodiversität 2017 im Naturpark Südsteiermark. Mitteilungen des Naturwissenschaftlichen Vereines für Steiermark 147: 99–134.
- Laciny, A. 2021. Among the shapeshifters: parasite-induced morphologies in ants (Hymenoptera, Formicidae) and their relevance within the EcoEvoDevo framework. EvoDevo 12, 2 (doi:10.1186/s13227-021-00173-2).
- Lapeva-Gjonova, A., Antonova, V. 2022. An updated checklist of ants (Hymenoptera, Formicidae) of Bulgaria, after 130 years of research. Biodiversity Data Journal 10, e95599 (doi:10.3897/bdj.10.e95599).
- Larabee, F.J., Suarez, A.V. 2014. The evolution and functional morphology of trap-jaw ants (Hymenoptera: Formicidae). Myrmecological News 20: 25-36.
- Latreille, P. A. 1802b. Histoire naturelle générale et particulière des Crustacés et des insectes. Tome 3. Familles naturelles des genres. Paris: F. Dufart, xii + 467 pp. (page 248, queen, male described)
- Latreille, P. A. 1804. Tableau méthodique des insectes. Pp. 129-200 in: Société de Naturalistes et d'Agriculteurs. Nouveau dictionnaire d'histoire naturelle. Tome 24. Paris: Déterville, 84 + 85 + 238 + 18 + 34 pp. (page 179, Combination in Myrmica)
- Leclerc, J.-B., Detrain, C. 2017. Loss of attraction for social cues leads to fungal-infected Myrmica rubra ants withdrawing from the nest. Animal Behaviour 129, 133-141 (DOI 10.1016/j.anbehav.2017.05.002).
- Lehue, M., Collignon, B., Detrain, C. 2020. Multiple nest entrances alter foraging and information transfer in ants. Royal Society Open Science 7, 191330 (doi:10.1098/RSOS.191330).
- Leppänen, J., Seppä, P., Vepsäläinen, K., Savolainen, R. 2016. Mating isolation between the ant Myrmica rubra and its microgynous social parasite. Insectes Sociaux 63: 79-86 (DOI 10.1007/s00040-015-0438-y).
- Leppänen, J., Vepsäläinen, K., & Savolainen, R. 2011. Phylogeography of the ant Myrmica rubra and its inquiline social parasite. Ecology and Evolution 1, 46–62 (doi:10.1002/ece3.6).
- Linnaeus, C. 1758. Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus I. Editio decima, reformata. Holmiae [= Stockholm]: L. Salvii, 824 pp. (page 580, worker described)
- Lucas, H. 1853. Essai sur les animaux articules qui habitent l'ile de Crete. Revue et Magasin de Zoologie Pure et Appliquée 5(2): 487-491.
- Maák, I., Roelandt, G., d'Ettorre, P. 2020. A small number of workers with specific personality traits perform tool use in ants. eLife 9, e61298 (doi:10.7554/elife.61298).
- Malagocka, J., Eilenberg, J., Jensen, A.B. 2019. Social immunity behaviour among ants infected by specialist and generalist fungi. Current Opinion in Insect Science 33, 99–104 (doi:10.1016/j.cois.2019.05.001).
- Maziarz, M., Broughton, R.K., Casacci, L.P., Dubiec, A., Maák, I., Witek, M. 2020. Thermal ecosystem engineering by songbirds promotes a symbiotic relationship with ants. Scientific Reports 10, 20330 (doi:10.1038/s41598-020-77360-z).
- Meadley-Dunphy, S.A., Prior, K.M. & Frederickson, M.E. 2019. Invasive ants disperse seeds farther than native ants, affecting the spatial pattern of seedling recruitment and survival. Oecologia doi:10.1007/s00442-019-04553-3.
- Meurville, M.-P., LeBoeuf, A.C. 2021. Trophallaxis: the functions and evolution of social fluid exchange in ant colonies (Hymenoptera: Formicidae). Myrmecological News 31: 1-30 (doi:10.25849/MYRMECOL.NEWS_031:001).
- Mitra, A. 2013. Function of the Dufour’s gland in solitary and social Hymenoptera. Journal of Hymenoptera Research 35, 33–58 (doi:10.3897/jhr.35.4783).
- Mokadam, C. 2021. Native and non-native ant impacts on native fungi (M.A. thesis, Buffalo State University).
- Muir, D. 1954. Ants Myrmica rubra L. and M. scabrinodis Nylander as intermediate hosts of a cestode. Nature 173: 688-689 (doi:10.1038/173688b0).
- Naumann, K. and R. J. Higgins. 2015. The European fire ant (Hymenoptera: Formicidae) as an invasive species: impact on local ant species and other epigaeic arthropods. Canadian Entomologist. 147:592-601. doi:10.4039/tce.2014.69
- Naumann, K., M. Moniz De Sa, E. Lewis, and R. Noronha. 2017. Supercolonies of the invasive ant, Myrmica rubra (Hymenoptera: Formicidae) in British Columbia, Canada. Journal of the Entomological Society of British Columbia. 114:56-64.
- Nemet, E., Czekes, Z., Tausan, I., Marko, B. 2012. Contribution to the knowledge of the myrmecofauna of the Cefa Nature Park (North-Western Romania). Acta Scientiarum Transylvanica Biologia 20, 61-72.
- Novgorodova, T. 2021. Preventing transmission of lethal disease: Removal behaviour of Lasius fuliginosus (Hymenoptera: Formicidae) Towards Fungus Contaminated Aphids. Insects 12, 99. (doi:10.3390/insects12020099).
- Novgorodova, T. A. 2015b. Organization of honeydew collection by foragers of different species of ants (Hymenoptera: Formicidae): Effect of colony size and species specificity. European Journal of Entomology. 112:688-697. doi:10.14411/eje.2015.077
- Novgorodova, T.A., Biryukova, O.B. 2011. Some ethological aspects of the trophobiotic interrelations between ants (Hymenoptera: Formicidae) and larvae of the sawfly Blasticotoma filiceti (Hymenoptera: Blasticotomidae), European Journal of Entomology 108, 47-52.
- Onoyama, K. 1989a. Confirmation of the occurrence of Myrmica rubra (Hymenoptera: Formicidae) in Japan with taxonomic and ecological notes. Jpn. J. Entomol. 57: 131-135 (page 131, see also)
- Oswalt, D.A. 2007. Nesting and foraging characteristics of the black carpenter ant Camponotus pennsylvanicus DeGeer (Hymenoptera: Formicidae). Ph.D. thesis, Clemson University.
- Pearson, B. 1981. The electrophoretic determination of Myrmica rubra microgynes as a social parasite: possible significance in the evolution of ant social parasites. Pp. 75-84 in: Howse, P. E., Clement, J.-L. (eds.) Biosystematics of social insects. Systematics Association Special Volume No. 19. London: Academic Press, 346 pp. (page 75, see also)
- Phillips, A.L., Attewell, P. J. 2011. An occurrence of Lasius sabularum (Bondroit, 1918) and Lasius umbratus (Nylander, 1846) within the same nest in Lincolnshire, England. BWARS Newsletter Autumn 2011:22-24.
- Purkart, A., Kollár, J., Goffová, K. 2019. Fauna of ants (Hymenoptera: Formicidae) of selected sand habitats in Podunajsko Region, Slovakia. Naturae Tutela 23(1): 101-111.
- Radchenko, A. 2023. Myrmica ants (Hymenoptera: Formicidae) in Late Eocene European ambers. Annales Zoologici 73(4), 629-648 (doi:10.3161/00034541anz2023.73.4.006).
- Radchenko, A. G.; Czechowski, W.; Czechowska, W. 1997. The genus Myrmica Latr. (Hymenoptera, Formicidae) in Poland - a survey of species and a key for their identification. Ann. Zool. (Warsaw) 47: 481-500 (page 483, Senior synonym of europaea)
- Radchenko, A.G. & Elmes, G.W. 2010. Myrmica ants of the Old World. Fauna Mundi 3: 1-789.
- Radchenko, A.G., Fisher, B.L., Esteves, F.A., Martynova, E.V., Bazhenova, T.N., Lasarenko, S.N. 2023. Ant type specimens (Hymenoptera, Formicidae) in the collection of Volodymyr Opanasovych Karawajew. Communication 1. Dorylinae, Poneromorpha and Pseudomyrmecinae. Zootaxa, 5244(1), 1–32 (doi:10.11646/zootaxa.5244.1.1).
- Reznikova, Z. 2020. Spatial cognition in the context of foraging styles and information transfer in ants. Animal Cognition. (doi:10.1007/s10071-020-01423-x).
- Salata, S., Borowiec, L., Trichas, A. 2020. Review of ants (Hymenoptera: Formicidae) of Crete, with keys to species determination and zoogeographical remarks. Monographs of the Upper Silesian Museum No 12: 5–296 (doi:10.5281/ZENODO.3738001).
- Savolainen, R., Vepsalainen, K. 2003. Sympatric speciation through intraspecific social parasitism. Proceedings of the National Academy of Sciences 100(12): 7169–7174.
- Schar, S. & Nash, D.R. 2014. Evidence that microgynes of Myrmica rubra ants are social parasites that attack old host colonies. Journal of Evolutionary Biology, doi:10.1111/jeb.12482.
- Schär, S., Vila, R., Menchetti, M. 2023. First record of the introduced ant Myrmica specioides in the eastern United States. Sociobiology 70(4), e9153 (doi:10.13102/sociobiology.v70i4.9153).
- Schifani, E., Castracani, C., Spotti, F.A., Giannetti, D., Ghizzoni, M., Gobbi, M., Lencioni, V., Pedrotti, L., Grasso, D.A., Mori, A. 2021. Social parasite ants in the Alps: a new site of the vulnerable Myrmica myrmicoxena and new uppermost altitudinal limit for M. microrubra. Sociobiology, 68(4): e7276 (doi:10.13102/sociobiology.v68i4.7276).
- Schultner, E., Pulliainen, U. 2020. Brood recognition and discrimination in ants. Insectes Sociaux 67, 11–34 (doi:10.1007/s00040-019-00747-3).
- Seifert, B. 1988b. A taxonomic revision of the Myrmica species of Europe, Asia Minor, and Caucasia (Hymenoptera, Formicidae). Abh. Ber. Naturkundemus. Görlitz 62(3): 1-75 (page 5, Senior synonym of laevinodis (and its junior synonyms bruesi, champlaini and longiscapus))
- Seifert, B. 2024. Pleistocene population differentiation in the ant Myrmica scabrinodis (Hymenoptera: Formicidae) – a taxonomic borderline case. Soil Organisms 96(1): 11-21 (doi:10.25674/357).
- Shrestha B, Tanaka E, Hyun MW, Han JG, Kim CS, Jo JW, Han SK, Oh J, Sung JM, Sung GH. 2017. Mycosphere Essay 19. Cordyceps species parasitizing hymenopteran and hemipteran insects. Mycosphere 8(9): 1424–1442 (DOI 10.5943/mycosphere/8/9/8).
- Siddiqui, J. A., Li, J., Zou, X., Bodlah, I., Huang, X. 2019. Meta-analysis of the global diversity and spatial patterns of aphid-ant mutualistic relationships. Applied Ecology and Environmental Research 17: 5471-5524 (doi:10.15666/aeer/1703_54715524).
- Siddiqui, J.A., Bamisile, B.S., Khan, M.M., Islam, W., Hafeez, M., Bodlah, I., Xu, Y. 2021. Impact of invasive ant species on native fauna across similar habitats under global environmental changes. Environmental Science and Pollution Research 28(39), 54362–54382 (doi:10.1007/s11356-021-15961-5).
- Silva, P.S., Koch, E.B. de A., Arnhold, A., Delabie, J.H.C. 2022. Review of distribution modeling in ant (Hymenoptera: Formicidae) biogeographic studies. Sociobiology 69(4), e7775 (doi:10.13102/sociobiology.v69i4.7775).
- Smith, D. R. 1979. Superfamily Formicoidea. Pp. 1323-1467 in: Krombein, K. V., Hurd, P. D., Smith, D. R., Burks, B. D. (eds.) Catalog of Hymenoptera in America north of Mexico. Volume 2. Apocrita (Aculeata). Washington, D.C.: Smithsonian Institution Press, pp. i-xvi, 1199-2209. (page 1350, see also)
- Snegovaya, N., Shigayev, C. 2021. A checklist of the ants (Insecta, Formicidae) of Azerbaijan Republic. Iranian Journal of Animal Biosystematics 17(2): 179-207 (doi:10.22067/ijab.2022.67343.1000).
- Sondej, I., Domisch, T. 2024. Impact of large-scale fire and habitat type on ant nest density and species abundance in Biebrza National Park, Poland. Forests 151, 123 (doi:10.3390/f15010123).
- Stukalyuk, S., Radchenko, A., Akhmedov, A., Reshetov, A., Netsvetov, M. 2021. Acquisition of invasive traits in ant, Crematogaster subdentata Mayr (Hymenoptera: Formicidae) in urban environments. Serangga 26: 1-29.
- Stukalyuk, S., Radchenko, Y., Gonchar, O., Akhmedov, A., Stelia, V., Reshetov, A., Shymanskyi, A. 2021. Mixed colonies of Lasius umbratus and Lasius fuliginosus (Hymenoptera, Formicidae): when superparasitism may potentially develop into coexistence: a case study in Ukraine and Moldova. Halteres 12, 25–48 (doi:10.5281/zenodo.5753121).
- Stukalyuk, S.V., Kozyr, M.S., Netsvetov, M.V., Zhuravlev, V.V. 2020. Effect of the invasive phanerophytes and associated aphids on the ant (Hymenoptera, Formicidae) assemblages. Halteres 11: 56-89 (doi:10.5281/ZENODO.4192900).
- Stukalyuk, S.V., Radchenko, A., Reshetov, A., Akhmedov, A., Goncharenko, I. 2021. Comparative analysis of the population structure of Crematogaster subdentata and Lasius neglectus in the primary and secondary ranges (Hymenoptera: Formicidae). Fragmenta Entomologica 53, 43-51 (doi:10.13133/2284-4880/436).
- Tartally, A., Somogyi, A.Á., Révész, T., Nash, D.R. 2020. Host ant change of a socially parasitic butterfly (Phengaris alcon) through host nest take-over. Insects 11, 556 (doi:10.3390/INSECTS11090556).
- Tausan, I., Dauber, J., Trica, M.R., Marko, B. 2017. Succession in ant communities (Hymenoptera: Formicidae) in deciduous forest clear-cuts - an Eastern European case study. European Journal of Entomology 114, 92–100 (doi:10.14411/eje.2017.013).
- Trible, W., Kronauer, D.J.C. 2017. Caste development and evolution in ants: it's all about size. Journal of Experimental Biology 220, 53–62 (doi:10.1242/jeb.145292).
- Tseng, S.-P. 2020. Evolutionary history of a global invasive ant, Paratrechina longicornis (Dissertation_全文 ). Ph.D. thesis, Kyoto University.
- Vepsalainen, K., Ebsen, J.R., Savolainen, R., Boomsma, J.J. 2009. Genetic differentiation between the ant Myrmica rubra and its microgynous social parasite. Insectes Sociaux 56:425–437.
- Warren II, R.J., Bayba, S., Krupp, K.T. 2018. Interacting effects of urbanization and coastal gradients on ant thermal responses. Journal of Urban Ecology 4: 1-11 (doi:10.1093/jue/juy026).
- Waters, J.S., Keough, N.W., Burt, J., Eckel, J.D., Hutchinson, T., Ewanchuk, J., Rock, M., Markert, J.A., Axen, H.J., Gregg, D. 2022. Survey of ants (Hymenoptera, Formicidae) in the city of Providence (Rhode Island, United States) and a new northern-most record for Brachyponera chinensis (Emery, 1895). Check List 18(6), 1347–1368 (doi:10.15560/18.6.1347).
- Wegnez, P. 2017. Découverte de Myrmica lobicornis Nylander, 1846 et Lasius jensi Seifert, 1982, deux nouvelles espèces pour le Grand-Duché de Luxembourg (Hymenoptera: Formicidae). Bulletin de la Société royale belge d’Entomologie153, 46–49.
- Wetterer, J.K., Radchenko, A.G. 2011. Worldwide spread of the ruby ant, Myrmica rubra (Hymenoptera: Formicidae). Myrmecological News 14: 87-96.
- Wheeler, G. C.; Wheeler, J. 1953a [1952]. The ant larvae of the myrmicine tribe Myrmicini. Psyche (Camb.) 59: 105-125 (page 118, larva described)
- Wiezik, M., Svitok, M., Wieziková, A., Dovčiak, M. 2013. Shrub encroachment alters composition and diversity of ant communities in abandoned grasslands of western Carpathians. Biodiversity and Conservation 22, 2305–2320 (doi:10.1007/s10531-013-0446-z).
- Yarrow, I. H. H. 1955c. The type species of the ant genus Myrmica Latreille. Proc. R. Entomol. Soc. Lond. Ser. B 24: 113-115 (page 114, Senior synonym of laevinodis (and its junior synonyms bruesi, champlaini and longiscapus))
- Zhong, Y., Huang, Y., Wu, T., Liu, Z. 2024. A new spineless Myrmica species from China, with the reinstatement of M. zhengi Ma & Xu (Hymenoptera: Formicidae). Journal of Asia-Pacific Entomology 27, 102261 (doi:10.1016/j.aspen.2024.102261).
- Zhu, W., Wu, L., Duan, L., Xu, S. 2022. A checklist of ants (Hymenoptera: Formicidae) in northern Shaanxi Province, China, with one new species of genus Proformica Ruzsky, 1902, Journal of Asia-Pacific Entomology 25, 101875 (doi:10.1016/j.aspen.2022.101875).
References based on Global Ant Biodiversity Informatics
- Agosti D. 1983. Zur Insektenfauna der Umgebung der Vogelwarte Sempach, Kanton Luzern. XIII. Hymenoptera 2: Formicidae (Ameisen). Entomologische Berichte Luzern 10: 91-92.
- Agosti, D. and C.A. Collingwood. 1987. A provisional list of the Balkan ants (Hym. Formicidae) and a key to the worker caste. I. Synonymic list. Mitteilungen der Schweizerischen Entomologischen Gesellschaft, 60: 51-62
- Alvarado M., and L. Galle. 2000. Ant assemblages associated with lowland forests in the southern part of the great Hungarian plain. Acta Zoologica Academiae Scientarum Hungaricae 46(2): 79-102.
- AntArea. Accessed on February 5th 2014 at http://antarea.fr/fourmi/
- Antonov I. A. 2012. Ant complexes of Baikalsk town. The Bulletin of Irkutsk State University 4: 143-146.
- Antonov I. A. 2013. Ant Assemblages (Hymenoptera: Formicidae) of Cities of the Temperate Zone of Eurasia. Russian Journal of Ecology 44(6): 523526.
- Antonova V., and L. Penev. 2008. Classification of assemblages of ants in the green areas in Sofia City. Acta Zoologica Bulgarica 60(2): 103-110.
- ArtDatabanken Bugs (via GBIG)
- Asociacion Iberica de Mirmecologia. 2011. List of species collected during the Taxomara Lisboa 2011. Iberomyrmex 3: 30-31.
- Assing V. 1989. Die Ameisenfauna (Hym.: Formicidae) nordwestdeutscher Calluna-Heiden. Drosera 89: 49-62.
- Astruc C., J. F. Julien, C. Errard, and A. Lenoir. 2004. Phylogeny of ants based on morphology and DNA sequence data. Molecular Phylogenetics and Evolution 31: 880-893.
- Babik H. 2011. Ants of Botanical and Zoological Gardens of Warsaw (Poland). Entomologica romanica 16: 53.
- Babik H., C. Czechowski, T. Wlodarczyk, and M. Sterzynska. 2009. How does a strip of clearing affect the forest community of ants (Hymenoptera: Formicidae)? Fragmenta Faunistica 52(2): 125-141?
- Baroni Urbani C., and C. A. Collingwood. 1976. A Numerical Analysis of the Distribution of British Formicidae (Hymenoptera, Aculeata). Verhandlungen der Naturforschenden Gesellschaft in Basel 85: 51-91.
- Baroni Urbani C., and C. A. Collingwood. 1977. The zoogeography of ants (Hymenoptera, Formicidae) in Northern Europe. Acta Zoologica Fennica 152: 1-34.
- Barrett K. E. 1967. Ants in South Brittany. Entomologist's Record and Journal of Variation 79:112-116.
- Barrett K. E. J. 1968. Ants in western France. Entomologist 101: 153-155.
- Barrett K. E. J. 1968b. The distribution of ants in central southern England. Transactions of the Society for British Entomology 17: 235-250.
- Barrett K. E. J. 1970. Ants in France, 1968-69. Entomologist 103: 270-274.
- Baugnee J. Y. 2003. Camponotus piceus (Leach, 1825), fourmi nouvelle pour la faune belge decouverte dans le parc naturel Viroin-Hermeton (Hymenoptera: Formicidae). Bulletin S. R. B. E./K. B. V. E. 139: 219-225.
- Behr D., S. Lippke, and K. Colln. 1996. Zur kenntnis der ameisen von Koln (Hymenoptera, Formicidae). Decheniana-Beihefte (Bonn) 35: 215-232.
- Behr D., and K. Colln. 1993. Zur ameisenfauna (Hymenoptera, Formicidae) von Gonnersdorf (Kr. Daun). Dendrocopos 20: 148-160.
- Belskaya E., A. Gilev, and E. Belskii. 2017. Ant (Hymenoptera, Formicidae) diversity along a pollution gradient near the Middle Ural Copper Smelter, Russia. Environ Sci Pollut Res DOI 10.1007/s11356-017-8736-8
- Bernadou A., G. Latil, V. Fourcassié, and X. Espadaler. 2006. Etude des communautés de fourmis d'une vallée andorrane. Union International pour l'Etude des Insectes Sociaux, Colloque annuel de la section francaise, 4pp.
- Bernadou A., V. Fourcassié, and X. Espadaler. 2013. A preliminary checklist of the ants (Hymenoptera, Formicidae) of Andorra. Zookeys 277: 13-23.
- Bernadou A., X. Espadaler, A. Le Goff, and V. Fourcassie. 2015. Ant community organization along elevational gradients in a temperate ecosystem. Insect. Soc. 62:5971
- Bernadou, A., G. Latil, V. Fourcassié, and X. Espadaler. "Les formigues de la Vall del Madriu-Perafita-Claror : diversitat i distribució." Hàbitats, 13 (2006): 10-21.
- Bernard F. 1967. Faune de l'Europe et du Bassin Méditerranéen. 3. Les fourmis (Hymenoptera Formicidae) d'Europe occidentale et septentrionale. Paris: Masson, 411 pp.
- Blacker N. C. 1989. The ants (Hymenoptera, Formicidae) of the Gower Peninsula, West Glamorgan, South Wales. Entomologist's Record and Journal of Variation 101: 261-266.
- Blacker N. C. and C. A. Collingwood. 2002. Some significant new records of ants (Hymenoptera: Formicidae) from the Salisbury area, south Wiltshire, England, with a key to the British species of Lasius. British Journal of Entomology and Natural History 15: 25-46
- Blatrix R., C. Lebas, C. Galkowski, P. Wegnez, P. Pimenta, and D. Morichon. 2016. Vegetation cover and elevation drive diversity and composition of ant communities (Hymenoptera: Formicidae) in a Mediterranean ecosystem. – Myrmecological News 22: 119-127.
- Boer P. 2019. Species list of the Netherlands. Accessed on January 22 2019 at http://www.nlmieren.nl/websitepages/specieslist.html
- Boer P., W. Dekoninck, A. J. Van Loon, and F. Vankerkhoven. 2003. Lijst van mieren (Hymenoptera: Formicidae) van Belgie en Nederland, hun Nederlandse namen en hun voorkomen. Entomologische Berichten (Amsterdam) 63: 54-58.
- Boer P., W. Dekoninck, A. J. van Loon, and F. Vankerkhoven. 2003. Lijst van mieren (Hymenoptera: Formicidae) van Belgie en Nederland, hun Nederlandse namen en hun voorkomen. Entomologische Berichten 63(3): 54-57.
- Boer P., W. Dekoninck, A. J. van Loon, and F. Vankerkhoven. 2003. List of ants (Hymenoptera: Formicidae) of Belgium and The Netherlands, their status and Dutch vernacular names. Entomologische Berichten 63 (3): 54-58.
- Boevé, J.-L. and F.L. Wäckers. 2003. Gustatory Perception and Metabolic Utilization of Sugars by Myrmica rubra Ant Workers. Oecologia 136(4):508-514
- Bonaric J. C. 1971. Contribution a l'etude systematique et ecologique des formicides du Bas-Languedoc. PhD thesis Universite des sciences et techniques du Languedoc, 175 pages.
- Bonaric J. C. 1971. Étude systématique et écologique des fourmis de lHérault. Ann. Soc. Hortic. Hist. Nat. Hérault 111: 81-87.
- Borowiec L. 2014. Catalogue of ants of Europe, the Mediterranean Basin and adjacent regions (Hymenoptera: Formicidae). Genus (Wroclaw) 25(1-2): 1-340.
- Borowiec L., and S. Salata. 2012. Ants of Greece - Checklist, comments and new faunistic data (Hymenoptera: Formicidae). Genus 23(4): 461-563.
- Boven J. K. A. 1947. Liste de détermination des principales espèces de fourmis belges (Hymenoptera Formicidae). Bulletin et Annales de la Société Entomologique de Belgique 83: 163-190.
- Boven J. van 1949. Notes sur la faune des Hautes-Fagnes en Belgique. Bulletin et Annales de la Société Entomologique de Belgique 85: 135-143.
- Bracko G. 2007. Checklist of the ants of Slovenia (Hymenoptera: Formicidae). Natura Sloveniae 9: 15-24
- Bracko G., H. C. Wagner, A. Schulz, E. Gioahim, J. Maticic, and A. Tratnik. 2014. New investigation and a revised checklist of the ants (Hymenoptera: Formicidae) of the Republic of Macedonia. North-Western Journal of Zoology 10(1): 10-24.
- Bracko, G. 2006. Review of the ant fauna (Hymenoptera:Formicidae) of Croatia. Acta Entomologica Slovenica 14(2): 131-156.
- Bracko, G. "Review of the ant fauna (Hymenoptera: Formicidae) of Croatia." Acta Entomologica Slovenica Vol 14 st (2006): 131-156.
- Brangham A. N. 1938. Additions to the wild fauna and flora of the Royal Botanic gardens, Kew: XVIII. Bulletin of Miscellaneous Information (Royal Botanic Gardens, Kew), 9: 390-396.
- Braschler, B. and B. Baur. 2005. Experimental Small-Scale Grassland Fragmentation Alters Competitive Interactions among Ant Species. Oecologia 143(2):291-300
- Bujan J., A. Brigic, Z. Sedlar, and R. Sostaric. 2015. Progressive vegetation succession of fen habitats promotes the lack of habitat specialist ants. Insectes Sociaux 62: 415-422.
- Carniel A. 1998. Ricerche sulla mirmecofauna delle Prealpi Orobiche (Lombardia) (Insecta, Hymenoptera, Formicidae). Atti. Mus. Civ. Stor. Nat. Morbegno 9: 29-39.
- Casevitz-Weulersse J. 1990. Etude Systematique de la Myrmecofaune Corse (Hymenoptera: Formicidae), deuxieme partie. Bull. Mus. Natn. Hist. Nat. Paris. 4eme serie 12, section A(2): 415-442.
- Casevitz-Weulersse J., and C. Galkowski. 2009. Liste actualisee des Fourmis de France (Hymenoptera, Formicidae). Bull. Soc. Entomol. Fr. 114: 475-510.
- Casevitz-Weulersse J., and M. Prost. 1991. Fourmis de la Côte-d'Or présentes dans les collections du Muséum d'Histoire Naturelle de Dijon. Bulletin Scientifique de Bourgogne 44: 53-72.
- Chen Ying-Wu et al. 2008. Preliminary list of Formicidae in Gansu Province. Journal of Anhui Agri. Sci. 36(32): 14133-14134
- Cherix D., and S. Higashi. 1979. Distribution verticale des fourmis dans le Jura vaudois et recensement prelimaire des bourdons (Hymenoptera, Formicidae et Apidae). Bull. Soc. Vaud. Sc. Nat. 356(74): 315-324.
- Colindre L. 2015. Les fourmis en Picardie: bilan 2014 (Hymenoptera/ Formicidae). Entomologiste Picard 26, 15 pages.
- Colindre L. 2017. Richess et utilite du cortege de fourmis en foret d'Ermenonville, Oise, Region Hauts-de-France. Association des Entomologistes de Picardie. 19 pages.
- Collingwood C. A. 1951. The distribution of ants in north-west Scotland. Scottish Naturalist 63: 45-49
- Collingwood C. A. 1951. The distribution of ants in north-west Scotland. Scottish Naturalist 63: 45-49.
- Collingwood C. A. 1955. Ants in S.W. Scotland. Entomologist's Record and Journal of Variation 67: 11-12.
- Collingwood C. A. 1956. Ant hunting in France. Entomologist 89: 106-108.
- Collingwood C. A. 1961. Ants in the Scottish Highlands. Scotish Naturalist 70: 12-21.
- Collingwood C. A. 1962. Some ants (Hym. Formicidae) from north-east Asia. Entomologisk Tidskrift 83: 215-230.
- Collingwood C. A. 1971. A synopsis of the Formicidae of north Europe. Entomologist 104: 150-176
- Collingwood C. A., and J. E. Satchell. 1956. The ants of the South Lake District. Journal of the Society for British Entomology 5: 159-164.
- Collingwood C. A., and J. Hughes. 1987. Ant species in Yorkshire, England. Naturalist (Leeds) 112: 95-101.
- Collingwood C., and A. Prince. 1998. A guide to ants of continental Portugal (Hymenoptera: Formicidae). Boletim da Sociedade Portuguesa de Entomologia. Suplemento 5: 1-49.
- Collingwood C.A. 1955. Ants in S.W. Scotland. Entomol.Rec. 67: 11-12
- Collingwood C.A. 1959. Ants in the Scottish Highlands. The Scottish Naturalist. 70: 12-21
- Collingwood C.A. 1959. Scandinavian Ants. Entomol. Rec. 71: 78-83
- Collingwood C.A. 1961. Ants in Finland. Entomol. Rec. 73: 190-195
- Collingwood C.A. 1961. New Vice-County Records for British Ants. Entomologist. 73: 90-93
- Collingwood C.A. and Satchell J.E. 1956. The Ants of the South Lake District. Journal of the Society for British Entomology. 5: 159-164
- Collingwood, C. A. 1958. The ants of the genus Myrmica in Britain. Proc. R. Entomol. Soc. Lond. Ser. A 33: 65-75
- Collingwood, C. A. 1958b. A key to the species of ants (Hymenoptera, Formicidae) found in Britain. Trans. Soc. Br. Entomol. 13: 69-96
- Collingwood, C. A. 1964. The Identification of British Ants (Hym. Formicidae). Transactions of the Society for British Entomology. 16:93-121.
- Collingwood, C. A. 1974. A revised list of Norwegian ants (Hymenoptera: Formicidae). Norsk Entomologisk Tidsskrift 21: 31-35.
- Collingwood, C. A., and I. H. H. Yarrow. "A survey of Iberian Formicidae." EOS (Revista española de entomología) 44 (1969): 53-101.
- Collingwood, C. A. "The Formicidae (Hymenoptera) of Fennoscandia and Denmark." Fauna Entomologica Scandinavica 8 (1979): 1-174.
- Collingwood, C.A. 1958. A survey of Irish Formicidae. Proceedings of the Royal Irish Academy 59B:213-219
- Consani M., and P. Zangheri. 1952. Fauna di Romagna. Imenotteri - Formicidi. Memorie della Societa Entomologica Italiana 31: 38-48.
- Csosz S., B. Marko, K. Kiss, A. Tartally, and L. Galle. 2002. The ant fauna of the Ferto-Hansag National Park (Hymenoptera: Formicidae). In: Mahunka, S. (Ed.): The fauna of the Fert?-Hanság National Park. Hungarian Natural History Museum, Budapest, pp. 617-629.
- Csősz S., B. Markó, and L. Gallé. 2001. Ants (Hymenoptera: Formicidae) of Stana Valley (Romania): Evaluation of the effectiveness of a myrmecological survey. Entomologica Romanica 6 : 121-126.
- Csősz S., B. Markó, and L. Gallé. 2011. The myrmecofauna (Hymenoptera: Formicidae) of Hungary: an updated checklist. North-Western Journal of Zoology 7: 55-62.
- Czechowski W., A. Radchenko, W. Czechowska and K. Vepsäläinen. 2012. The ants of Poland with reference to the myrmecofauna of Europe. Fauna Poloniae 4. Warsaw: Natura Optima Dux Foundation, 1-496 pp
- Czechowski W., M. Woyciechowski, and W. Czechowska. 1999. Myrmica microrubra Seifert, 1993 (Hymenoptera, Formicidae) - an inquiline ant species new to Poland. Fragmenta Faunistica 42: 123-126.
- Czekes Z., Radchenko, A. G., Csősz, S. Szász-Len, A., Tăuşan, I., Benedek, K., and Markó, B. 2013. The genus Myrmica Latreille, 1804 (Hymenoptera: Formicidae) in Romania: distribution of species and key for their identification. Entomologica Romanica 17: 29-50.
- Dauber J. 1997. Ameisenfauna einer urbanen landschaft. Naturschutz und Landschaftsplanung 29: 303-309.
- Dauber, J., J. Bengtsson and L. Lenoir. 2006. Evaluating Effects of Habitat Loss and Land-Use Continuity on Ant Species Richness in Seminatural Grassland Remnants. Conservation Biology 20(4):1150-1160
- Dekoninck W., H. De Koninck, J. Y. Baugnee, and J. P. Maelfait. 2007. Ant biodiversity conservation in Belgian calcareous grasslands: active management is vital. Belg. J. Zool. 137 (2): 137-146.
- Dekoninck W., K. Desender, and P. Grootaert. 2008. Establishment of ant communities in forests growing on former agricultural fields: Colonisation and 25 years of management are not enough (Hymenoptera: Formicidae). Eur. J. Entomol. 105: 681689.
- Della Santa E. 1995. Fourmis de Provence. Faune Provence 16: 5-37.
- Della Santa E. 2000. L'identification des espèces du genre Myrmica Latreille (Formicidae) de Suisse; essai de présentation synoptique. Bull. Romand Entomol. 18: 169-187.
- Dewes E. 2005. Ameisenerfassung im Waldschutzgebiet Steinbachtal/Netzbachtal. Abh. Delattinia 31: 89-118.
- Dietrich C. O., B. Schlick, and F. Steiner. 1998. Ameisen bei Hochwasser (Hymenoptera: Formicidae) Beobachtungen in Ostösterreich im Juli 1997. Myrmecologische Nachrichten 2: 35-41.
- Donisthorpe H. 1914. Myrmecophilous notes for 1913. Entomologist's Record and Journal of Variation 26: 37-45.
- Dubovikoff D. A., and Z. M. Yusupov. 2018. Family Formicidae - Ants. In Belokobylskij S. A. and A. S. Lelej: Annotated catalogue of the Hymenoptera of Russia. Proceedingss of the Zoological Institute of the Russian Academy of Sciences 6: 197-210.
- Dvorak, L., P. BOGUSCH, I. MALENOVSKÝ, P. BEZDÌÈKA, K. BEZDÌÈKOVÁ, K. HOLÝ, P. LIKA, J. MACEK, L. ROLLER, M. RÍHA et al. "Hymenoptera of Hády Hill, near the city of Brno (Czech Republic), collected during the Third Czech-Slovak Hymenoptera meeting." Acta Musei Moraviae, Scientiae biologicae (Brno) 93 (2008): 53-92.
- Ebsen J. R., J. J. Boomsma, and D. R. Nash. 2019. Phylogeography and cryptic speciation in the Myrmica scabrinodis Nylander, 1846 species complex (Hymenoptera: Formicidae), and their conservation implications. Insect Conservation and Diversity doi: 10.1111/icad.12366
- Elmes G. W. 1973. Miniature queens of the ant Myrmica rubra L. (Hymenoptera, Formicidae). Entomologist 106: 133-136.
- Elmes G. W. 1994. A population of the social parasite Myrmica hirsuta Elmes (Hymenoptera, Formicidae) recorded from Jutland, Denmark, with a first description of the worker caste. Insectes Soc. 41: 437-442.
- Elmes, G.W., T. Akino, J.A. Thomas, R.T. Clarke and J.J. Knapp. 2002. Interspecific differences in cuticular hydrocarbon profiles of Myrmica ants are sufficiently consistent to explain host specificity by Maculinea (large blue) butterflies. Oecologia 130:525-535
- Else G., B. Bolton, and G. Broad. 2016. Checklist of British and Irish Hymenoptera - aculeates (Apoidea, Chrysidoidea and Vespoidea). Biodiversity Data Journal 4: e8050. doi: 10.3897/BDJ.4.e8050
- Emery C. 1914. Wissenschaftliche Ergebnisse der Bearbeitung von O. Leonhard's Sammlungen. 5. Südeuropäische Ameisen (Hym.). Entomologische Mitteilungen. Berlin-Dahlem 3: 156-159.
- Emery C. 1916. Fauna entomologica italiana. I. Hymenoptera.-Formicidae. Bullettino della Società Entomologica Italiana 47: 79-275.
- Emery, C. "Catalogo delle formiche esistenti nelle collezioni del Museo Civico di Genova. Parte seconda. Formiche dell'Europa e delle regioni limitrofe in Africa e in Asia." Annali del Museo Civico di Storia Naturale 12 (1878): 43-59.
- Entomological Society of Latvia. 2003. http://leb.daba.lv/Formicidae.htm (Accessed on December 1st 2013).
- Espadaler X., F. Garcia, X. Roig, and R. Vila. 2013. Ants (Hymenoptera, Formicidae) from the Castell de Montesquiu park (Osona, north-east of the Iberian Peninsula). Boletín de la Sociedad Entomológica Aragonesa (S.E.A.) 53: 223-227.
- Espadaler X., X. Roig, K. Gómez, and F. García. 2011. Formigues de les Planes de Son i mata de València (Hymenoptera, Formicidae) Treballs de la Institució Catalana d'Història Natural 16: 609-627.
- Espadaler, X., and X. Roig. "Ants from the Montnegre-Corredor Natural Park with description of the male lasius cinereus Seifert." Miscellanea Zoologica 23 (2) (2001): 45-53. Abstract
- Espadaler, X. "Contribución al conocimiento de los formícidos (Hymenoptera, Formicidae) del Pirineo catalán." Tesis Universida (1979): 285 pp.
- Fiedler, K., F. Kuhlmann, B. C. Schlick-Steiner, F. M. Steiner and G. Gebauer. 2007. Stable N-isotope signatures of central European ants assessing positions in a trophic gradient. Insectes Sociaux 54(4):393-402.
- Finzi B. 1926. Le forme europee del genere Myrmica Latr. Primo contributo. Bollettino della Società Adriatica di Scienze Naturali in Trieste. 29: 71-119.
- Forel A. 1892. Die Ameisenfauna Bulgariens. (Nebst biologischen Beobachtungen.). 305-318.
- Forel A. 1901. Variétés myrmécologiques. Annales de la Société Entomologique de Belgique 45: 334-382.
- Forel A. 1906. Les fourmis de l'Himalaya. Bulletin de la Société Vaudoise des Sciences Naturelles 42: 79-94.
- Forel A. 1911. Sur le genre Metapone n. g. nouveau groupe des Formicides et sur quelques autres formes nouvelles. Rev. Suisse Zool. 19: 445-459.
- Formidabel Database
- Fowles, A.P. 1996. A provisional checklist of the invertebrates recorded from Wales. 2. Aculeate wasps, bees and ants (Hymenoptera: Aculeata). Countryside Council for Wales
- Francoeur A., and R. Béique. 1966. Les Formicides (Hyménoptères) de Provancher. Can. Entomol. 98: 140-145.
- Freitag A. 2013. Biodiversite 2010 en ville de Neuchatel: les fourmis (Hymenopteres, Formicidae). Bulletin de la societe Neuchateloise des sciences naturelles 133: 183-200.
- GRETIA. 2017. Bilan annuel de l'enquete sur la repartition des fourmis armoricaines. 23 pages.
- Gadeau de Kerville H. 1922. Materiaux pour la Faune des Hymenopteres de la Normandie. Bull. Soc. Amis Sc. Nat. Rouen 1916-1921, 1922: 217-225.
- Galkowski C. 2013. Nouvelles données sur la répartition de Strongylognathus huberi Forel, 1874 (Hymenoptera, Formicidae) en France. Bulletin de la Société Linnéenne de Bordeaux (n.s.) 41: 167-174.
- Galkowski C., and C. Foin. 2013. Nouvelles données sur la répartition de Strongylognathus huberi Forel, 1874 (Hymenoptera, Formicidae) en France. Bulletin de la Société Linnéenne de Bordeaux (n.s.) 41: 167-174.
- Galkowski C., and P Wegnez. 2010. Myrmica constricta Karavaiev 1934, nouvelle espece pour la France (Hymenoptera, Formicidae). Bull. Soc. Ent. Mulhouse 66(3): 41-45.
- Galle L. 1981. The Formicoid fauna of the Hortobagy. Pp. 307-311 in: Mahunka, S. (ed.) 1981. The fauna of the Hortobágy National Park. Budapest: Akadémiai Kiadó, 415 pp.
- Galle L. 1993. Data to the ant fauna of the Bukk (Hymenoptera: Formicidae). Natural history of the national parks of Hungary 7: 445-448.
- Galle L. 1997. Contribution to the ant fauna of Slovenia with special reference to the submediterranean and eudinaric regions. Annals for Istrian and Mediterranean studies 11: 209-214.
- Gallé L. 1991. Structure and succession of ant assemblages in a north European sand dune area. Holarctic Ecology 14: 31-37.
- Gallé L., B. Markó, K. Kiss, E. Kovács, H. Dürgő, K. Kőváry, and S. Csősz. 2005. Ant fauna of Tisza river basin (Hymenoptera: Formicidae). In: Gallé, L. (szerk.): Vegetation and Fauna of Tisza River Basin I. Tiscia Monograph Series 7; Szeged, pp. 149-197.
- Garcia Garcia F., and A. D. Cuesta-Esgura. 2017. First catalogue of the ants of Burgos province, Spain (Hymenoptera: Formicidae). Boletín de la Sociedad Entomológica Aragonesa 60: 245–258.
- García F., X. Espadaler, P. Echave, and R. Vila. 2011. Hormigas (Hymenoptera, Formicidae) de los acantilados de l'Avenc de Tavertet (Osona) Boletín de la Sociedad Entomológica Aragonesa 47: 363-367.
- Gaspar C. 1966. Etude myrmecologique des tourbieres dans les Hautes-Fagnes en Belgique (Hymenoptera, Formicidae). Rev. Ecol. Biol. Sol. 3(2): 301-312.
- Gaspar C. 1966. Etude myrmecologique des tourbieres dans les Huates-Fagnes en Belgique (Hymenoptera, Formicidae). Rev. Ecol. Biol. Sol. 3(2): 301-312.
- Gaspar C. 1968. Les fourmis de la Drome et des Basses-Alpes, en France (Hymenoptera, Formicidae). Naturaliste can. 95: 747-766.
- Gaspar C., and C. Thirion. 1978. Modification des populations d'Hymenopteres sociaux dans les milieux anthropogenes. Memorabilia Zoologica 29: 61-77.
- Gaspare Charles. 1965. Étude myrmécologique d'une région naturelle de Belgique: la Famenne Survey des Fourmis de la Région (Hymenoptera, Formicidae). Institut agronomique de l'Etat a' Gembloux. 32(4): 427-434.
- Gibb H., and T. Johansson. 2011. Field tests of interspecific competition in ant assemblages: revisiting the dominant red wood ants. Journal of Animal Ecology 80: 548-557.
- Gilev A. V., I. V. Kuzmin, V. A. Stolbov, and S. D. Sheikin. 2012. Materials on the fauna and ecology of ants (formicidae) Southern part of the Tyumen region. Tyumen State University Herald 6: 86-91.
- Glaser F. 2009. Die Ameisen des Fürstentums Liechtenstein. (Hymenoptera, Formicidae). Amtlicher Lehrmittelverlag, Vaduz, 2009 (Naturkundliche Forschung im Fürstentum Liechtenstein; Bd. 26).
- Glaser F., A. Freitag, and H. Martz. 2012. Ants (Hymenoptera: Formicidae) in the Münstertal (Val Müstair) a hot spot of regional species richness between Italy and Switzerland. Gredleriana 12: 273 - 284.
- Glaser F., T. Kopf, and K. H. Steiberger. 2003. Ameisen (Hymenoptera: Formicidae) im Frastanzer Ried und den Illauen (Vorarlberg, Österreich) Artenspektrum, Gefährdung und Schutzempfehlungen. Vorarlberger Naturschau 13: 287-310.
- Gouraud C. 2015. Bilan de l’année 2014 : Atlas des fourmis de Loire-Atlantique (Hymenoptera, Formicidae). Atlas des Formicidae de Loire-Atlantique, compte rendu de la première année d’étude (2014)
- Grandi G. 1935. Contributi alla conoscenza degli Imenotteri Aculeati. XV. Boll. R. Ist. Entomol. Univ. Studi Bologna 8: 27-121.
- Gratiashvili N., Barjadze S. 2008. Checklist of the ants (Formicidae Latreille, 1809) of Georgia. Proceedings of the Institute of Zoology (Tbilisi) 23: 130-146.
- Grzes I. M. 2009. Ant species richness and evenness increase along a metal pollution gradient in the Boles?aw zinc smelter area. Pedobiologia 53: 65-73.
- Grzes I. M., P. Slipinski, H. Babik, D. Moron, B. Walter, G. Trigos Peral, I. Maak, and M. Witek. 2018. Colony size and brood investment of Myrmica rubra ant colonies in habitats invaded by goldenrods. Insectes Sociaux 65:275–280.
- Gulzar A. 2014. Classification and Distribution of ants in Kashgar, Xinjiang. Master's Thesis Shaanxi Normal University, 75 pages.
- Guénard B., and R. R. Dunn. 2012. A checklist of the ants of China. Zootaxa 3558: 1-77.
- Hauschteck-Jungen E., and H. Jungen. 1983. Ant chromosomes. II. Karyotypes of western palearctic species. Insectes Soc. 30: 149-164.
- Heinken, T. 2004. Migration of an Annual Myrmecochore: A Four Year Experiment with Melampyrum pratense L. Plant Ecology 170(1):55-72
- Hicks B. J., B. L. Pilgrim, and H. D. Marshall. 2014. Origins and genetic composition of the European fire ant (Hymenoptera: Formicidae) in Newfoundland, Canada. Canadian Entomologist 146(4): 457-464.
- Holgersen H. 1940. Myrmekologiske notiser I. Nor. Entomol. Tidsskr. 5: 183-187.
- Holgersen H. 1942. Ants of northern Norway (Hym., Form.). Tromso Mus. Årsh. 63(2): 1-34.
- Holgersen H. 1943. Ant studies in Rogaland (south-western Norway). Avhandlingar utgitt av det Norske Videnskaps-Akademi i Oslo. I. Matematisk-Naturvidenskapelig Klasse 1943(7): 1-75.
- Holgersen H. 1944. The ants of Norway (Hymenoptera, Formicidae). Nytt Magasin for Naturvidenskapene 84: 165-203.
- Holgersen H. 1944. The ants of Norway (Hymenoptera, Formicidae). Nytt Magasin for Naturvidenskapene 84: 165-488.
- Holgersen H. 1944. The ants of Norway (Hymenoptera, Formicidae). Nytt Magasin for Naturvidenskapene 84: 165-518.
- Hölzel E. 1952. Ameisen Kärntens. Carinthia II 142: 89-132.
- IZIKO South Africa Museum Collection
- Ihnatiuk O. A., and S. V. Stukalyuk. 2015. Degradation changes in the structure of multispecies associations of ants in urbanized areas. Russian Journal of Ecology 46(1): 109–115.
- Jakubzik A., H. Kinkler, and K. Colln. 2010. Aculeate Hymenoptera from a Humid Biotope in Leverkusen-Steinbüchel. Decheniana (Bonn) 163: 145158.
- Jansen G., R. Savolainen, K. Vespalainen. 2010. Phylogeny, divergence-time estimation, biogeography and social parasite–host relationships of the Holarctic ant genusMyrmica(Hymenoptera: Formicidae). Molecular Phylogenetics and Evolution 56: 294-304.
- Jeffery H. G. 1931. The Formicidae (or ants) of the Isle of Wight. Proceedings of the Isle of Wight Natural History and Archaeological Society 2: 125-128.
- Karaman M. G. 2011. A catalogue of the ants (Hymenoptera, Formicidae) of Montenegro. Podgorica: Catalogues 3, Volume 2, Montenegrin Academy of Sciences and Arts, 140 pp.
- Kiran K., and C. Karaman. 2012. First annotated checklist of the ant fauna of Turkey (Hymenoptera: Formicidae). Zootaxa 3548: 1-38.
- Kiseleva E. F. 1925. On the ant fauna of the Ussuri region. Izv. Tomsk. Gos. Univ. 75: 73-75.
- Kiss K., and K. Fetyko. 2008. Notes about the ant fauna (Hymenoptera: Formicidae) of the Tur valley. In. Sike T., Márk-Nagy J. (eds), The flora and fauna of the Tur river natural reserve, Biharean Biologist, II, Oradea, pp.71-76.
- Kofler A. 1995. Nachtrag zur Ameisenfauna Osttirols (Tirol, Österreich) (Hymenoptera: Formicidae). Myrmecologische Nachrichten 1: 14-25.
- Korlevic, A. "Prilozi fauni hrvatskih opnokrilaca." Glasn. Hrv. Narav. Dr. 5 (1890): 189-250.
- Kozisek T. 1987. Ants (Formicoidea) of the Abrod State Nature Reserve. Ochrana Prírody 8: 205-208.
- Krugova T. M. 2017. Ant's population in the chern taiga of Tigireksky mountain range (North-Western Altai). Proceedings of the Tigirek Reserve 9: 68-77.
- Kupianskaya A. N., Lelej, A.S., and Urbain, B. K. 2000. The Ants (Hymenoptera: Formicidae) of the Kuril Islands. Far Eastern Entomologist. 92:1-21.
- Kupianskaya A. N., and A. S. Lelej. 2000. The ants (Hymenoptera, Formicidae) collected in the Habomai and Shikotan (Kuril Islands) in 1998. Far Eastern entomologist 92: 22-24.
- Kupianskaya, A. N., Lelej, A.S., and Urbain, B. K. 2000. The Ants (Hymenoptera: Formicidae) of the Kuril Islands. Far Eastern Entomologist. 92:1-21.
- Kvamme T. 1982. Atlas of the Formicidae of Norway (Hymenoptera: Aculeata). Insecta Norvegiae 2: 1-56.
- Kvamme T. 1999. Notes on Norwegian ants (Hymenoptera, Formicidae). Norwegian Journal of Entomology 46: 19-23.
- Kvamme T., and A. Wetas. 2010. Revidert liste over norske maur Inkludert dialektiske navn og forslag til nye norske navn og forslag til norske navn. Norsk institutt for skog og landskap, Ås. 127 pp
- Laeger T., and R. Schultz. 2005. Ameisen (Hymenoptera: Formicidae) als Beifänge in Bodenfallen wie genau spiegeln sie reale Abundanzverhältnisse wider? Myrmecologische Nachrichten 7: 17-24.
- Lapeva-Gjonova, L., V. Antonova, A. G. Radchenko, and M. Atanasova. "Catalogue of the ants (Hymenoptera, Formicidae) of Bulgaria." ZooKeys 62 (2010): 1-124.
- Le Moli F., and A. Zaccone. 1995. Ricerche sulla mirmecofauna del Cansiglio (Prealpi Carniche). Soc. Ven. Sc. Nat. 20: 33-52.
- Lebas C., C. Galkowski, P. Wegnez, X. Espadaler, and R. Blatrix. 2015. The exceptional diversity of ants on mount Coronat (Pyrénées-Orientales), and Temnothorax gredosi(Hymenoptera, Formicidae) new to France. R.A.R.E., T. XXIV (1): 24 33
- Legakis Collection Database
- Lehouck V., D. Bonte, W. Dekoninck, and J. P. Maelfait. 2004. The distribution of ant nests (Hymenoptera, Formicidae) in coastal grey dunes of Flanders (Belgium) and their relationship to myrmecochorous plants. Belg. J. Zool. 134 (2/1) : 89-96.
- Lenoir A. 1971. Les fourmis de Touraine, leur intérêt biogéographique. Cahiers des Naturalistes 27: 21-30.
- Li Z.h. 2006. List of Chinese Insects. Volume 4. Sun Yat-sen University Press
- Lillig M., and E. Dewes. 2015. The former Siegfried Line as habitats for ants (Hymenoptera: Formicidae). Abh. Delattinia 37: 117 - 136
- Liu C., K. L. Dudley, Z. Xu, and E. P. Economo. 2017. Mountain metacommunities: climate and spatial connectivity shape ant diversity in a complex landscape. Ecography 10.1111/ecog.03067.
- Liu X. 2012. Taxonomy, diversity and spatial distribution characters of the ant family Formicidae (Insecta: Hymenoptera) in southeastern Tibet. PhD Thesis 139 pages
- Liu X., Z. H. Xu, X. Y. Zhou, N. N. Yu, and C. L. Zhang. 2011. Ant communities of West slope of Mount Demula and Bomi Valley in Southeastern Tibet. Forest Research 24(4): 458-463.
- Liu X., Z. Xu, C. Zhang, N. Yu, and G. Xu. 2012. Distribution patterns of ants from West slope of Mount Demula and Bomi Valley in Southeastern Tibet. Journal of Northwest Forestry University 27(4): 77-82.
- Liu X., Z. Xu, X. Zhou, N. Yu, and C. Zhang. 2011. Ant Communities of West Slope of Mount Demula and Bomi Valley in Southeastern Tibet. Forest Research 24(4): 458-463.
- Livory A. 2003. Les fourmis de la Manche. L'Argiope 39: 25-49.
- Lomnicki J. 1928. Spis mrówek Lwowa i okolicy. Ksiegi Pamiatkowej (Lecia Gimn. IV Jana Dlugosza Lwowie) 50: 1-10.
- Lorite P., J. A. Carrillo, M. F. García, and T. Palomeque. 2000. Chromosome numbers in Spanish Formicidae. III. Subfamily Myrmicinae (Hymenoptera). Sociobiology 36: 555-570.
- Maavara V. 1953. Ants of Estonian SSR. ABIKS loodusevaatlejale 10: 1-44.
- Majzlan O., and P. Devan. 2009. Selected insect groups (Hymenoptera, Neuroptera, Mecoptera, Raphidioptera) of the Rokoš Massif (Strážovské vrchy Mts.). Rosalia (Nitra), 20, p. 63–70.
- Malozemova L. A. 1972. Ants of steppe forests, their distribution by habitats, and perspectives of their utilization for protection of forests (north Kazakhstan). [In Russian.]. Zoologicheskii Zhurnal 51: 57-68.
- Marko B. 1999. Contribution to the knowledge of the myrmecofauna of the River Somes valley. In: Sárkány-Kiss, A., Hamar, J. (szerk.): The Somes/Szamos River Valley. A study of the geography, hydrobiology and ecology of the river system and its environment. Tiscia Monograph Series 3, Szolnok-Szeged-Tîrgu Mure?, pp. 297-302.
- Marko B. 2008. Ants (Hymenoptera: Formicidae) of the Sand Dunes of Foieni Protected Area and Its Surroundings (Satu Mare County, Romania), and a New Species for the Romanian Fauna. Acta Scientiarum Transylvanica 16(3): 87-99.
- Marko B., K. Kiss, and L. Galle. 2004. Mosaic structure of ant communities (Hymenoptera: Formicidae) in Eastern Carpathian marshes: regional versus local scales. Acta Zoologica Academiae Scientiarum Hungaricae 50(2): 7795.
- Markó B. 1997. Contribution to the Knowledge of the Ant-Fauna (Hymenoptera: Fromicoidea) of the Cri?ul-Repede River-Valley. In: Sárkány-Kiss, A., Hamar, J. (szerk.): The Cri?/Körös Rivers' Valleys. A study of the geography, hydrobiology and ecology of the river system and its environment. Tiscia Monograph Series 2, Szolnok-Szeged-Tîrgu Mure?, pp. 345-352.
- Markó B., B. Sipos, S. Csősz, K. Kiss, I. Boros, and L. Gallé. 2006. A comprehensive list of the ants of Romania (Hymenoptera: Formicidae). Myrmecologische Nachrichten 9: 65-76.
- Markó B., and S. Csősz. 2002. Die europäischen Ameisenarten (Hymenoptera: Formicidae) des Hermannstädter (Sibiu, Rumänien) Naturkundemuseums I.: Unterfamilien Ponerinae, Myrmicinae und Dolichoderinae. Annales Historico-Naturales Musei Nationalis Hungarici 94: 109-121.
- Martorell, M. "Catálogos sinonímicos de insectos encontrados en Cataluña." Barcelona (1879).
- Menozzi C. 1918. Primo contributo alla conoscenza della fauna mirmecologica del Modenese. Atti della Società dei Naturalisti e Matematici di Modena. (5)4: 81-88.
- Meyer M., C. Braunert, R. Gerend, and I. Schrankel. 2003. Résultats de l'excursion annuelle du groupe de travail entomologique de la Société des naturalistes luxembourgeois. Bull. Soc. Nat. luxemb. 103: 103-120.
- Monteserín Real, S. "Invertebrados de la Reserva Natural Integral de Muniellos, Asturias: Formicidae." Consejería de Medio Ambiente, Ordenación del Territorio e Infraestructuras del Principado de Asturias KRK edicio (2003): 269.
- Moscaliuc L. 2008. Notes on the ant fauna (Hymenoptera: Formicidae) of Rarau mountain. Analele Facult??ii de Biologie, Univ A.I. Cuza Ia?i 54: 53-55.
- Mouquet C. 2006. Premier inventaire des invertébrés terrestres des plages du département du Calvados. Rapport Gretia pour le Syndicat Mixte Calvados Littoral Espaces Naturels. 36 p.
- Nadig A. 1918. Alcune note sulla fauna dell'alta Valsesia. Formicidae. Atti Soc. Ital. Sci. Nat. Mus. Civ. Stor. Nat. Milano 56: 331-341.
- Nemet E., Z. Czekes, I. Tausan, and B. Marko. 2012. Contribution to the knowledge of the myrmecofauna of the Cefa Nature Park (North-Western Romania). Acta Scientiarum Transylvanica 20(1): 61-72.
- Neumeyer R., and B. Seifert. 2005. Commented check list of free living ant (Hymenoptera: Formicidae) species of Switzerland. Bulletin de la Societe Entomologique Suisse 78: 1-17.
- Novgorodova T. A., A. S. Ryabinin. 2015. Trophobiotic associations between ants (Hymenoptera, Formicidae) and aphids (Hemiptera, Aphidomorpha) in South Zauralye. News of Saratov University. Chemistry Series, Biology, Ecology 2(15): 98-107.
- Nylander, W. "Synopsis des Formicides de France et d'Algérie." Annales des Sciences Naturelles, Zoologie (4)5 (1856): 51-109.
- O'Rourke F. J. 1948. The distribution and general ecology of the Irish Formicidae. Proceedings of the Royal Irish Academy. Section B: Biological, Geological, and Chemical Science 52: 383-410.
- Onoyama, K. 1989a. Confirmation of the occurrence of Myrmica rubra (Hymenoptera: Formicidae) in Japan with taxonomic and ecological notes. Japanese Journal of Entomology 57:131-135. [1989-03-25] PDF 127635
- Onoyama, K. 1989a. Confirmation of the occurrence of Myrmica rubra (Hymenoptera: Formicidae) in Japan with taxonomic and ecological notes. Japanese Journal of Entomology 57:132. [1989-03-25] PDF 127635
- Ottonetti L., L. Tucci, and G. Santini. 2006. Recolonization Patterns of Ants in a Rehabilitated Lignite Mine in Central Italy: Potential for the Use of Mediterranean Ants as Indicators of Restoration Processes. Restoration Ecology 14(1): 6066.
- Paraschivescu D. 1972. Fauna mirmecologica din zonele saline ale Romaniei. Studii si Cercetari de Biologie. Seria Zoologie 24: 489-495.
- Paraschivescu D. 1978. Elemente balcanice in mirmecofauna R. S. Romania. Nymphaea 6: 463- 474.
- Paukkunen J., O. Bistrom, E. Budrys, E. Helve, C. Lagercrantz, I. Mannerkosko, S. Orlovskyte, and M. Tahtinen. 2017. Entomological excursion to the Curonian Spit in August 2016. Naujos ir Retos Lietuvos Vabzdziu Rusys 28: 97-120.
- Paukkunen J., and M. V. Kozlov. 2015. Stinging wasps, ants and bees (Hy menoptera: Aculeata) of the Murmansk region, Northwest Russia. — Entomol. Fennica. 26: 53–73.
- Petal J. 1974. Analysis of a sheep pasture ecosystem in the Pieniny mountains (the Carpathians) XV. The effect of pasture management on ant population. Ekologia Polska 22(3/4): 679-692.
- Petal J. 1980. The effect of industrial pollution of Silesia on populations of ants. Polish Ecological Studies 6(4): 665-672.
- Petal J. M. 1963. Faune des fourmis de la reserve de tourbiere en projet a Rakowskie Bagno pres de Frampol (voivodie de Lublin). Annales Universitatis Mariae Curie-Sk?odowska 58(7): 143-174.
- Petal J. M. 1967. Contribution a la connaissance des fourmis (Formicidae, Hymenoptera) de la region de Lublin. VII. Associations des fourmis des milieux de tourbieres, de forets et de dunes aux environs de Libiszow (dist. De Parczew). Annales Universitatis Mariae Curie-Sklodowska Lublin-Polonia 22(9): 117-130.
- Petal J. M. 1976. The effect of mineral fertilization on ant populations in meadows. Polish Ecological Studies 2(4): 209-218.
- Petal J., H. Jakubczyk, and Z. Wojcik. 1970. Influence des fourmis sur les modifications des sols et des plantes dans les milieurs de prairie. In: Methods of study in soil ecology. Ed. J. Philipson. Pp. 235-239.
- Petal J., and A. Kusinska. 1994. Fractional composition of organic matter in the soil of anthills and of the environement of meadows. Pedobiologia 38: 493-501.
- Petrov I. Z. 2002. Contribution to the myrmecofauna (Formicidae, Hymenoptera) of the Banat Province (Serbia). Archives of Biological Sciences, Belgrade, 54(12): 57-64.
- Petrov I. Z. 2010. Contribution to the myrmecofauna (Hymenoptera: Formicidae) of Krusevac and its surroundings. Acta entomologica serbica, 2010, 15(1): 145-148.
- Petrov I. Z. 2012. Preliminary data on ants (Formicidae, Hymenoptera) on Mountain Avala (Belgrade, Serbia). Bulletin of the Natural History Museum 5: 95-99.
- Petrov I. Z., B. Petrov, D. Milicic, T. Karan-Znidarsic. 2007. Contribution to the Myrmecofauna (Hymenoptera: Formicidae) of East and South Serbia. Acta Zoologica Bulgarica 59(3): 295-299.
- Petrov I. Z., and C. A. Collingwood. 1992. Survey of the myrmecofauna (Formicidae, Hymenoptera) of Yugoslavia. Archives of Biological Sciences (Belgrade) 44: 79-91.
- Radchenko A. G., and G. W. Elmes. 2003. A taxonomic revision of the socially parasitic Myrmica ants (Hymenoptera: Formicidae) of the Palaearctic region. Annales Zoologici (Warsaw) 53: 217-243.
- Radchenko A. G., and G. W. Elmes. 2010. Myrmica ants (Hymenoptera: Formicidae) of the Old World. Fauna Mundi 3. Warsaw: Natura Optima Dux Foundation, 790 pp.
- Reznikova Z. I. 2003. Distribution patterns of ants in different natural zones and landscapes in Kazakhstan and West Siberia along a meridian trend. Euroasian Entomological Journal 2(4): 235-342.
- Ryabinin A. S., and T. A. Novgorodova. 2013. The ant fauna (Hymenoptera, Formicidae) of South Zauralye of Russia. Euroasian Entomological Journal 12(2): 161-166.
- Röszler P. 1950. Die Ameisenwelt des Nagy Pietrosz, 2305 m (Ungarn) und Umgebung. Zool. Anz. 145: 210-225.
- Saaristo M. I. 1995. Distribution maps of the outdoor myrmicid ants (Hymenoptera, Formicidae) of Finland, with notes on their taxonomy and ecology. Entomol. Fennica 6: 153-162.
- Sadil J. V. 1952. A revision of the Czechoslovak forms of the genus Myrmica Latr. (Hym.). Sb. Entomol. Oddel. Nár. Mus. Praze 27: 233-278.
- Salata S. 2014. Ants (Hymenoptera: Formicidae) of the National Park of the Stołowe Mts. Przyroda Sudetow 17: 161-172.
- Sanchez-Gil Jimeno R., and J. L. Reyes-Lopez. 2016. Study of ants species of the Sierra de San Carlos del Valle (Ciudad Real) and updating the provincial check list (Hymenoptera, Formicidae). Boln. Asoc. esp. Ent. 40 (1-2): 93-109.
- Santschi F. 1931. Notes sur le genre Myrmica (Latreille). Revue Suisse de Zoologie 38: 335-355.
- Santschi, F. "Liste de fourmis d'Espagne recueilliés par Mr. J. M. Dusmet." Boletín de la Sociedad Entomológica de España 15 (1932): 69-74.
- Schar S., G Talavera, X. Espadaler, J. D. Rana, A. A. Andersen, S. P. Cover, and R. Vila. 2018. Do Holarctic ant species exist? Trans-Beringian dispersal and homoplasy in the Formicidae. Journal of Biogeography 00: 1-12.
- Schlick-Steiner B. C., F. M. Steiner, G. Bracko, W. Paill, and B. Seifert. 2003. The temporary social parasite Lasius bicornis (Fórster, 1850) new to Slovenia, and Myrmica bessarabica Nassonov, 1889 corrected into M. specioides Bondroit, 1918 (Hymenoptera: Formicidae). Acta Entomol. Slov. 11: 159-162.
- Schlick-Steiner B. C., and F. M. Steiner. 1999. Faunistisch-ökologische Untersuchungen an den freilebenden Ameisen (Hymenoptera: Formicidae) Wiens. Myrmecologische Nachrichten 3: 9-53.
- Schultz, R., A. G. Radchenko, and B. Seifert. "A critical checklist of the ants of Kyrgyzstan (Hymenoptera: Formicidae)." Myrmecologische Nachrichten 8 (2006): 201-207.
- Seifert B. 1988. A taxonomic revision of the Myrmica species of Europe, Asia Minor, and Caucasia (Hymenoptera, Formicidae). Abhandlungen und Berichte des Naturkundemuseums Görlitz 62(3): 1-75.
- Seifert B. 1993. Taxonomic description of Myrmica microrubra n sp. - a social parasitic ant so far known as the microgyne of Myrmica rubra (L.). Abhandlungen und Berichte des Naturkundemuseums Görlitz 67(5): 9-12.
- Seifert B. 1994. Die freilebenden Ameisenarten Deutschlands (Hymenoptera: Formicidae) und Angaben zu deren Taxonomie und Verbreitung. Abhandlungen und Berichte des Naturkundemuseums Görlitz 67(3): 1-44.
- Slipinski P., B. Marko, K. Rzeszowski, H. Babik, and W. Czechowski. 2014. Lasius fuliginosus(Hymenoptera: Formicidae) shapes local ant assemblages. North-Western Journal of Zoology 10(2): 404-412.
- Slipinski P., M. Zmihorski, and W. Czechowski. 2012. Species diversity and nestedness of ant assemblages in an urban environment. Eur. J. Entomol. 109: 197206.
- Sonnenburg H. 2005. Die Ameisenfauna (Hymenoptera: Formicidae) Niedersachsens und Bremens. Braunschweiger Naturkundliche Schriften 7: 377-441.
- Soulie J. 1962. Fourmis des Hautes-Pyrenees. Bull. Soc. Hist. Nat. Toulouse 97: 35-37.
- Soyer B. 1951. Les fourmis des collines des environs de Marseille. Bull Soc Linn Provence 18: 3-6.
- Steffek J., and M. Wiezik. 2008. Exploited peatbog and associated mollusk and ant assemblages: still a reasonable protection? Naturae Tutela 12: 15-19.
- Steiner F. M., B. C. Schlick-Steiner, H. Konrad, K. Moder, E. Christian, B. Seifert, R. H. Crozier, C. Stauffer, and A. Buschinger. 2006. No sympatric speciation here: multiple data sources show that the ant Myrmica microrubra is not a separate species but an alternate reproductive morph of Myrmica rubra. Journal of Evolutionary Biology 19: 777-787.
- Steiner F. M., S. Schödl, and B. C. Schlick-Steiner. 2002. Liste der Ameisen Österreichs (Hymenoptera: Formicidae), Stand Oktober 2002. Beiträge zur Entomofaunistik 3: 17-25.
- Stiprais M. 1973. Materi?li par R?gas kukai?u faunu. - Latvijas Entomologs, 15: 30-32.
- Stitz H. 1934. Schwedisch-chinesische wissenschaftliche Expedition nach den nordwestlichen Provinzen Chinas, unter Leitung von Dr. Sven Hedin und Prof. Sü Ping-chang. Insekten gesammelt vom schwedischen Arzt der Expedition Dr. David Hummel 1927-1930. 25. Hymenoptera. Ark. Zool. 27A(1) 11: 1-9.
- Stukalyuk S. V. 2015. Structure of the ant assemblages (Hymenoptera, Formicidae) in the broad-leaved forests of Kiev. Entomological Review 95(3): 370–387.
- Stukalyuk S. V. 2016. Crematogaster subdentata Mayr, 1877, a potentially invasive species of ant (Hymenoptera, Formicidae) new to the fauna of Crimea. Entomological Review 95(8)
- Stukalyuk S. V. 2017. Stratification of the ant species (Hymenoptera, Formicidae) in the urban broadleaf woodlands of the city of Kiev. Entomological Review 97(3): 320-343.
- Stumper R. 1953. Etudes myrmecologiques. XI. Fourmis luxembourgeoises. Bulletin Soc. Nat. luxemb. 57: 122-135.
- Sushko G. G. 2013. The ants (hymenoptera, formicidae) bogs Belarusian Lakeland. Proceedings of the international. scientific-practical. conference, Vitebsk, 21-22 November 2013 - Vitebsk, 2013. - P. 215-216.
- Suvak M. 2007. The ants (Hymenoptera: Formicidae) in the stubble-fields of east Slovakia. Acta Zoologica Universitatis Comenianae. 47: 21-33
- Suñer i Escriche, David. "Contribució al coneixement mirmecologic de Gavarres, Montgrí, Guilleríes i la Serralada Transversal." Tesis Doctoral Universida (1991): 577 pp.
- Sveum P. 1979. Notes on the distribution of some Norwegian ant species (Hymenoptera, Formicidae). Fauna Norv. ser. B. 26: 10-11.
- Szujecki A., J. Szyszko, S. Mazur, and S. Perlinski. 1978. A succession of the ants (Formicidae) on afforested arable land and forest soils. Memorabilia Zoologica 29: 183-189.
- Tartally A. 2009. Data on the ant fauna of Gy?r?f? (SW-Hungary) (Hymenoptera: Formicidae). Natura Somogyiensis 13: 155-158.
- Tausan I. 2009. Contribution to the knowledge of the myrmecofauna (Hymenoptera, Formicidae) of the upper Aries river Basin (Transylvania, Romania). Transylv. Rev. Syst. Ecol. Res. 7: 87-96.
- Tausan I. 2010. Notes on the ant fauna (Hymenoptera, Formicidae) of the Rodna Mountains National Park and it's surroundings (Transylvania-Maramures, Romania). Transylv. Rev. Syst. Ecol. Res. 9: 159-166.
- Tausan I., J. Dauber, M. R. Tricia, and B. Marko. 2017. Succession in ant communities (Hymenoptera: Formicidae) in deciduous forest clear-cuts – an Eastern European case study. European Journal of Entomology 114: 92-100.
- Tausan I., M. M. Jerpel, I. R. Puscasu, C. Sadeanu, R. E. Brutatu, L. A. Radutiu, and V. Giurescu. 2012. Ant fauna (Hymenoptera: Formicidae) of Sibiu County (Transylvania, Romania). Brukenthal. Acta Musei 7(3): 499-520.
- Tausan I., and B. Marko. 2009. Comparative analysis of ant communities (Hymenoptera: Formicidae) in the surroundings of Sibiu (Romania). Brukenthal. Acta Musei 4(3): 635-644.
- Tausan I., and I C. Negrila. 2015. The ant collection (Hymenoptera: Formicidae) of the natural history museum of Sibiu (Romania). Brukenthal Acta Musei 3: 409-418.
- Terayama M. 1992. Structure of ant communities in East Asia. A. Regional differences and species richness. Bulletin of the Bio-geographical Society of Japan 47: 1-31.
- Terayama M., K. Ogata, and B.M. Choi. 1994. Distribution records of ants in 47 prefectures of Japan. Ari (report of the Myrmecologists Society of Japan) 18: 5-17.
- Terayama. M. 2004. Geological and ecological distribution of Japanese ants communities. (translated from Japanese) Reports of the Saitama Prefecture Animal Research Association. 48:23
- Tie Ru, and Xu Shengquan. 2004. Variety and distribution of ants in Northwest China. Journal of Ningxia Agricultural College 25(3): 4-9.
- Urbieta Balado A., A. Palanca Soler, C. Castan Lanaspa, and X. Espadaler. 1984. Ants in the supraforestal pastures of the Tendenera massif. Documents dEcologie Pyrénéenne III-IV: 223-224.
- Vagalinski B., and A. Lapeva-Gjonova. 2012. The ants (Hymenoptera, Formicidae) of Plana Mountain (Bulgaria). Historia naturalis bulgarica 20: 87-101.
- Vele A., J. Holisa, J. Tragnerova. 2015. Ant succession on burned areas in forested landscape: a case study from the Bohemian Switzerland National Forest. Zpravy Lesnickeho Vyzkumu 60(1): 47-52.
- Vele A., J. Holusa, J. Horak. 2016. Ant abundance increases with clearing size. J For Res 21: 110–114.
- Vele A., J. Holusa, and J. Frouz. 2009. Sampling for ants in different-aged spruce forests: A comparison of methods. European Journal of Soil Biology 45(4): 1-6.
- Vepsalainen K., H. Ikonene, and M. J. Koivula. 2008. The structure of ant assembalges in an urban area of Helsinki, southern Finland. Ann. Zool. Fennici 45: 109-127.
- Vepsäläinen, K., J. R. Ebsen, R. Savolainen and J. J. Boomsma. 2009. Genetic differentiation between the ant Myrmica rubra and its microgynous social parasite. Insectes Sociaux 56(4):425-437.
- Vogrin, V. "Prilog fauni Hymenoptera - Aculeata Jugoslavije." Zast. Bilja 31(suppl.) (1955): 1-74.
- Weber N. A. 1947. A revision of the North American ants of the genus Myrmica Latreille with a synopsis of the Palearctic species. I. Annals of the Entomological Society of America 40: 437-474.
- Wegnez P. 2017. Découverte de Myrmica lobicornis Nylander, 1846 et Lasius jensi Seifert, 1982, deux nouvelles espèces pour le Grand-Duché de Luxembourg (Hymenoptera: Formicidae). Bulletin de la Société royale belge d’Entomologie 153: 46-49.
- Wegnez P. 2018. Premières decouvertes de Myrmica bibikoffi Kutter, 1963 et de Ponera testacea Emery, 1895, au Luxembourg (Hymenoptera: Formicidae)Bulletin de la Société royale belge d’Entomologie 154: 263–272.
- Wegnez P., D. Ignace, E. Lommelen, M. Hardy, J. Bogaert, and C. Nilsson. 2015. Redécouverte de Teleutomyrmex schneideriKutter, 1950 dans les Alpes françaises (Hymenoptera: Formicidae). Bulletin de la Société royale belge d’Entomologie 151: 52-57.
- Wegnez P., and A. Ronk. 2017. Découverte de Camponotus herculeanus (Linnaeus, 1758) et signalement de quelques autres espèces rares de fourmis au Luxembourg (Hymenoptera : Formicidae). Bulletin de la Société des naturalistes luxembourgeois 119 : 153–159.
- Wegnez P., and F. Mourey. 2016. Formica uralensis Ruzsky, 1895 une espèce encore présente en France mais pour combien de temps ? (Hymenoptera: Formicidae). Bulletin de la Société royale belge d’Entomologie 152: 72-80.
- Wegnez P., and M. Fichaux. 2015. Liste actualisee des especes de fourmis repertoriees au Grand-Duche de Luxembourg (Hymenoptera: Formicidae). Bulletin de la Société royale belge d’Entomologie 151: 150-165
- Wei C., S. Y. Zhou, H. He, and M. T. Liu. 2001. A taxonomic study of the genus Myrmica Latreille from China. (Hymenoptera: Formicidae). Acta Zootaxonomica Sinica 26: 560-564.
- Wei C., Zhou S.-Y. and He H. 2001. A taxonomic study of the genus Myrmica Latreille from China (Hymenoptera: Formicidae). Acta Zootaxonomica Sinica 26(4): 560-564.
- Wheeler W. M. 1906. The ants of Japan. Bulletin of the American Museum of Natural History 22: 301-328.
- Wiezik M. 2007. Ants (Hymenoptera: Formicidae) of mountain and alpine ecosystems at Southern part of Krá?ovoho?ské Tatry Mts. Naturae Tutela 11: 85-90.
- Wiezik M., A. Wiezikova, and J. Murin. 2008. Ant assemblages at horná orava peatbogs. Naturae Tutela 12: 55-60.
- Wiezik M., A. Wiezikova, and M. Svitok. 2010. Effects of secondary succession in abandoned grassland on the activity of ground-foraging ant assemblages (Hymenoptera: Formicidae). Acta Soc. Zool. Bohem. 74: 153-160
- Wiezik M., A. Wiezikova, and M. Svitok. 2011. Vegetation structure, ecological stability, and low-disturbance regime of abandoned dry grasslands support specific ant assemblages in Central Slovakia. Tuexenia 31: 301315.
- Wiezik M., A. Wieziková, and L. Ma?áková. 2010. Ant assemblages (Hymenoptera: Formicidae) of typical habitats at Boky National Nature Reserve. Naturae Tutela 14(1): 91-98.
- Wiezik M., M. Kozon, and A. Wiezikova. 2013. Ants of selected peat bog habitats at the Horná Orava protected area. Naturae Tutela 17(1): 57-63.
- Wieziková A. 2008. Epigaeic activity of ants (Hymenoptera: Formicidae) at hemerobic gradient: arable land green forage permanent grassland. Acta Facultatis Ecologiae 18: 69-76.
- Wlodarczyk T. 2010. Ant species composition in relation to forest associations in Szczecin Landscape Park. Polish Journal of Entomology 79: 91-99.
- Wu wei, Li Xiao Mei, Guo Hong. 2004. A primary study on the fauna of Formicidae in Urumqi and its vicinities. Arid Zone Research 21(2): 179-182
- Xin M., Y. Ma, and D. He. 2011. Fauna composition of Formicidae in Ningxia. Journal of Ningxia University (Natural Science Edition) 32(4): 403-412.
- Xu Z., J. Chu, C Zhang, and N. Yu. 2011. Ant species and distribution pattern in Gongbo Nature Reserve in Southeastern Tibet. Sichuan Journal of Zoology 30(1): 118-123.
- Xu Z., J. Chu, C. Zhang, and N. Yu. 2011. Ant Species and D istribution Pattern in GongboNature Reserve in Southeastern T ibet. Sichuan Journal of Zoology 30(1): 118-123.
- Yasuda M., and F. Koike. 2009. The contribution of the bark of isolated trees as habitat for ants in an urban landscape. Landscape and Urban Planning 92: 276-281.
- Yoshimura M. 2000. Ants in the island of Hokkaido (Part 3, Part ???) Ants from Islands in Hokkaido, Northern Japan (No. 3, Teuri Island). Rishiri Studies 19: 27-35.
- Zhang C., Z. Xu, N. Yu, Q. He, and X. Liu. 2012. Distribution Patterns of Ant Species on East Slope of Mount Demola and Zayu Valley in Southeastern Tibet. Journal of Northeast Forestry University 40(3): 87-92
- Zhuytszyuan D. 2016. The ants (Hymenoptera, Formicidae) Nizhne-Svirsky reserve and their environmental features. Master's thesis Saint Petersburg State University.
- Zryanin V. A., and T. A. Zryanina. 2007. New data on the ant fauna Hymenoptera, Formicidae in the middle Volga River Basin. Uspekhi Sovremennoi Biologii 127(2): 226-240.
- Pages using DynamicPageList3 parser function
- Common Name
- Invasive
- Polygynous
- Supercolonies
- Photo Gallery
- North temperate
- Ant Associate
- Host of Formica fusca
- Host of Lasius niger
- Host of Lasius platythorax
- Host of Leptothorax muscorum
- Pteromalid wasp Associate
- Host of Asaphes vulgaris
- Host of Pachyneuron aphidis
- Encyrtid wasp Associate
- Host of Syrphophagus aphidivorus
- Aphid Associate
- Host of Cinara apini
- Mite Associate
- Host of Petalomium podolicus
- Host of Scutacarus longisetus
- Nematode Associate
- Host of Mermithidae (unspec.)
- Host of Mermithidae (unspecified "Mermix")
- Cestode Associate
- Host of Davaineidae (Raillietina urogalli??)
- Host of Raillietina urogalli
- Gregarine Associate
- Host of Mattesia geminata
- Fungus Associate
- Host of Ophiocordyceps myrmecophila
- Host of Rickia wasmanniii
- Host of Ophiocordyceps kniphofioides
- Host of Ophiocordyceps myrmicarum
- Host of Ophiocordyceps subramanianii
- Host of Rickia wasmannii
- FlightMonth
- Karyotype
- Species
- Extant species
- Formicidae
- Myrmicinae
- Myrmicini
- Myrmica
- Myrmica rubra
- Myrmicinae species
- Myrmicini species
- Myrmica species
- Ssr