Myrmica sabuleti
| Myrmica sabuleti | |
|---|---|

| |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Arthropoda |
| Class: | Insecta |
| Order: | Hymenoptera |
| Family: | Formicidae |
| Subfamily: | Myrmicinae |
| Tribe: | Myrmicini |
| Genus: | Myrmica |
| Species group: | sabuleti |
| Species: | M. sabuleti |
| Binomial name | |
| Myrmica sabuleti Meinert, 1861
| |
| Synonyms | |
| |


{kind=link}
A host to the Large Blue butterfly (Phengaris arion). Note that the ‘‘West-Mediterranean” form of M. sabuleti (Seifert, 1988) is now known as Myrmica spinosior (Seifert 2005).
| At a Glance | • Polygynous |
Identification
Radchenko and Elmes (2010) - A member of the sabuleti complex of the scabrinodis species group. This complex contains all species with males having a relatively long scape (equal to total length of 4-4.5 basal funicular segments). The workers may be confused with several species, but generally they are characterized by a quite narrow frons (mean FI 0.33) and a well-developed lobe at the scape base. Males are distinguished from the socially parasitic members of this group, Myrmica hirsuta, Myrmica vandeli and Myrmica bibikoffi, by the absence of long erect to suberect hairs on the head margins.
Collingwood (1979) - Reddish brown. Antennal scapes are sharply angulate with a longitudinal keel running forward from the bend and a more or less massive lateral extension, which in Scandinavian samples is frequently curved up to appear as a large semiupright tooth seen from behind. The petiole node is more rounded and usually less truncate than in Myrmica scabrinodis Nyl. and the epinotal spines are relatively longer but these features are too variable for certain discrimination between the species in all cases. Head Index: 85.6; Frons Index: 36.8; Frontal Lamina Index: 66.5. Length: 4.0-5.0 mm.
Keys including this Species
- Key to Myrmica of West Europe and North Africa
- Key to Myrmica of East Europe, West Siberia, northern Kazakhstan, Caucasus, Asia Minor, Turkmenistan and Iran
- Key to Myrmica of Romania
- Key to European parasitic Myrmica queens
Distribution
Europe (to the north until southern Sweden, Finland and Norway), Caucasus; all previous records of this species east of Ural Mts. belong to M. lonae (or any other species in older papers).
This is a northern species recorded only from mainland provinces Epirus, Macedonia, Sterea Ellas, Thessaly and Thrace (Borowiec et al., 2022).
Latitudinal Distribution Pattern
Latitudinal Range: 65.256706° to 37.164722°.
| North Temperate |
North Subtropical |
Tropical | South Subtropical |
South Temperate |
- Source: AntMaps
Distribution based on Regional Taxon Lists
Palaearctic Region: Albania, Andorra, Austria, Balearic Islands, Belarus, Belgium, Bulgaria, Channel Islands, Croatia, Czechia, Denmark (type locality), Estonia, Finland, France, Germany, Greece, Hungary, Iberian Peninsula, Iran, Jersey, Latvia, Liechtenstein, Lithuania, Luxembourg, Montenegro, Netherlands, Norway, Poland, Portugal, North Macedonia, Republic of Moldova, Romania, Russian Federation, Slovakia, Slovenia, Spain, Sweden, Switzerland, Türkiye, Ukraine, United Kingdom of Great Britain and Northern Ireland.
Distribution based on AntMaps
Distribution based on AntWeb specimens
Check data from AntWeb
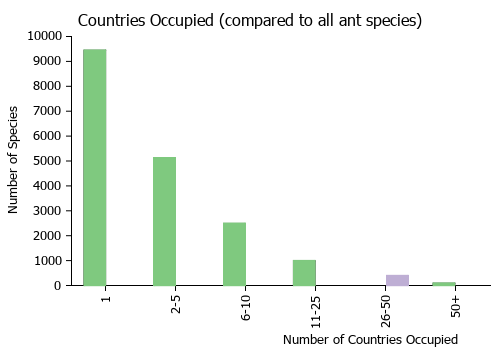
Countries Occupied
| Number of countries occupied by this species based on AntWiki Regional Taxon Lists. In general, fewer countries occupied indicates a narrower range, while more countries indicates a more widespread species. |
|
Biology
Radchenko and Elmes (2010) - The ecology of M. sabuleti has been relatively well studied during the last 20 years mainly because of its importance as the primary ant host of Phengaris arion, which figures in Red Data Books throughout Europe (Settele et al. 2005). The average size of nest is important in determining the suitability of a site to support a population of P. arion because Thomas and Wardlaw (1992) showed that it took the entire annual production of a nest of about 350 workers to rear one caterpillar. Also M. sabuleti has evolved interesing mutualisms with other grassland Lycaenid butterflies (e.g. Thomas 1983). It is also the host for several host-specific parasitic Myrmica species: Myrmica hirsuta, Myrmica bibikoffi and possibly Myrmica laurae as well as the more generalist Myrmica karavajevi. At this time it is not known why M. sabuleti should be such a good host for various social parasites. Perhaps it is related to the species’ habitat preferences or maybe to social factors, for example compared to other European Myrmica species, M. sabuleti workers generally lay far fewer eggs (Wardlaw and Elmes 1998); also it is somewhat easier to introduce foreign queens into existing cultures (personal observations) and it seems to have symbioses with aphids (see below).
For anyone who has studied their ecology it is hard to see how the female castes of M. sabuleti were ever confused with Myrmica scabrinodis: apart from the obvious morphological differences, they are adapted to living in warmer habitats than M. scabrinodis (e.g. Brian 1964; Brian et al. 1976; Doncaster 1983), both adults and larvae having a lower basal metabolism and larvae grow and develop more slowly than those of M. scabrinodis when reared at the same temperature (Elmes and Wardlaw 1983a) and workers have a longer foraging period (Elmes 1982). Habitats are usually grasslands, where M. sabuleti favour short-turf in northern latitudes and live under quite long grass in southern Europe (Thomas et a!. 1998). In Northwest Europe it could be considered a relict species of the post-glacial optimum that has persisted because of anthropogenic modifications of the natural environment (Thomas 1993). M. sabuleti is not normally considered a woodland species though it can live in open woodland glades, especially in eastern Europe, where summer temperatures are higher. It is often found in mature sand-dune systems, particularly those that have become grassed and are grazed by animals, the types were probably found in such a sandy place, hence its name.
Workers appear to be generalist predator scavengers, like most of the other Myrmica species. There is circumstantial evidence that it might eat the eliasome of Viola seeds. M. sabuleti do not often forage on obvious aphid colonies, more usually it maintains small cultures of aphids, particularly those species that live on the base of the stems at ground level; one can often find these colonies enclosed with soil and a few workers inside guarding and tending the aphids (personal observations). Workers were observed tending colonies of aphids on the underside of leaves of low-growing Clinopodium vulgare (Wood-Baker 1977) and we suspect that M. sabuleti may be more dependent upon such less-obvious associations than previously appreciated. Collingwood (1979) - This is a robust species usually nesting in sun exposed sheltered sites, often in groups of small nests each containing up to a 1000 or more workers with a few queens. It is characteristically larger and more brightly coloured than the similar Myrmica scabrinodis and easy to distinguish in Scandinavia where the scape development is relatively massive equivalent to the form described as var. lonae Finzi. Nests are usually located under stones but unlike M. scabrinodis seldom or never in tree stumps or in boggy land.
M. sabuleti mainly competes for living space with M. scabrinodis at the cooler end of its niche, and with Myrmica specioides and Myrmica schencki at the hotter end. The processes involved in these competitive interactions have been explored in numerical spatial-simulation models. The average numbers of workers per nest varied from 12 - > 4000 in a study of nearly 300 nests censused in southern England; average nest size on acid grassland was smaller (about 150 workers) than nests on chalk grassland (about 370 workers) but the average number of queens per nest in both types of habitat was the same (just over 1 per nest) (Elmes and Wardlaw, 1982a). A more detailed study of the chalk grassland population showed that above-nest vegetation was implicated in determining the size and productivity of the nest (Elmes and Wardlaw 1982b, c). A population living under stones on limestone grassland had much larger more polygynous colonies, averaging about 1200 workers with 15 queens per nest (Elmes 1974b), this was the population that contained the type series of the social parasite M. hirsuta (see Notes for that species above). Seppa (1996) demonstrated lowered levels of relatedness among workers commensurate with all queens actively reproducing. Brian (1972) followed the populations of several individual nests for some years and showed there to be quite a high turnover of queens and workers, one nest studied supported a population of the parasite Myrmica karavajevi. In an appendix to a paper Brian and Elmes (1974) showed that variations in the annual productivity of this population was directly related to changes in the annual insolation.
M. sabuleti has been used quite frequently in comparative studies of chemical ecology (e.g. Cammaerts et al. 1981; Evershed et. al. 1982). In a more recent study of the cuticular hydrocarbons (Elmes et al. 2002) it was shown that M. sabuleti had a rather “simpler” chemical profile than other species, in particular workers had less Dienes and less Di-methyl alcanes than M. scabrinodis, the two species can be easily discriminated by their chemical profiles. M. sabuleti has been used as a model species in studies of food competition and recruitment (De Vroey 1980a; Cammaerts and Cammaerts 1980; Debiseau and Pasteels 1994; Debiseau et al. 1997).
Nuptial flights occur from August to the end of September. Mixed swarms with other Myrmica species are common (Woyciechowski 1990c and personal observations) and where it co-exists with Myrmica lonae mixed flights are normal.
Collingwood (1979) - This is a robust species usually nesting in sun exposed sheltered sites, often in groups of small nests each containing up to a 1000 or more workers with a few queens. It is characteristically larger and more brightly coloured than the similar Myrmica scabrinodis and easy to distinguish in Scandinavia where the scape development is relatively massive equivalent to the form described as var. lonae Finzi. Nests are usually located under stones but unlike M. scabrinodis seldom or never in tree stumps or in boggy land.
Flight Period
| X | X | X | |||||||||
| Jan | Feb | Mar | Apr | May | Jun | Jul | Aug | Sep | Oct | Nov | Dec |
Source: antkeeping.info.
- Check details at Worldwide Ant Nuptial Flights Data, AntNupTracker and AntKeeping.
 Explore: Show all Flight Month data or Search these data. See also a list of all data tables or learn how data is managed.
Explore: Show all Flight Month data or Search these data. See also a list of all data tables or learn how data is managed.
Association with Other Organisms
- Explore: Show all Associate data or Search these data. See also a list of all data tables or learn how data is managed.
Other Insects
- This species is a host for the ant Myrmica bibikoffi (a temporary parasite) in Switzerland (de la Mora et al., 2021; Jansen et al., 2010).
- This species is a host for the ant Myrmica hirsuta (a inquiline) (de la Mora et al., 2021; Jansen et al., 2010).
- This species is a host for the ant Myrmica karavajevi (a workerless inquiline).
- This species is a host for the butterfly Maculinea arion (a parasite) (Barbero et al., 2009).
It is thought that this species is a host for the workerless inquiline Myrmica laurae, but this needs confirmation.
The record of this species hosting Myrmica lemasnei is incorrect and was a misidentification of Myrmica spinosior).
This ant has been associated with the butterflies Phengaris arion and a species that has recently been recognized as two distinct species: Polyommatus icarus and Polyommatus celin. Presently it is unclear if this latter association is between Myrmica sabuleti and one or the other of these species, or both (Obregon et al. 2015).
Nematode
- This species is a host for the nematode Mermithidae (unspec.) (a parasite) in Poland (Czechowsk et al., 2007; Laciny, 2021).
- This species is a host for the nematode Hexamermis/Agamermis (a parasite) in Germany (Steiner et al., 2019; Laciny, 2021).
Fungi
- This species is a host for the fungus Hormiscium myrmecophilum (a pathogen) (Espadaler & Santamaria, 2012).
This species is a host for the fungus Rickia wasmanniii (a pathogen) (Espadaler & Santamaria, 2012).
- This species is a host for the fungus Rickia wasmannii (a parasite) (Quevillon, 2018) (encounter mode primary; direct transmission; transmission within nest).
Life History Traits
- Queen number: polygynous (Rissing and Pollock, 1988; Frumhoff & Ward, 1992)
- Queen type: winged (Rissing and Pollock, 1988; Frumhoff & Ward, 1992) (queenless worker reproduction)
- Mean colony size: 3,000 (Brian, 1972; Cammaerts & Cammaerts, 1980; Beckers et al., 1989)
- Foraging behaviour: mass recruiter (Brian, 1972; Cammaerts & Cammaerts, 1980; Beckers et al., 1989)
Castes
Queen
Images from AntWeb
   
| |
| Queen (alate/dealate). Specimen code casent0173178. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by CAS, San Francisco, CA, USA. |
    
| |
| Queen (alate/dealate). Specimen code casent0173179. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by CAS, San Francisco, CA, USA. |
Nomenclature
The following information is derived from Barry Bolton's Online Catalogue of the Ants of the World.
- sabuleti. Myrmica sabuleti Meinert, 1861: 327 (w.m.) DENMARK. Emery, 1908a: 176 (q.); Hauschteck, 1965: 325 (k.); Donisthorpe, 1915b: 258 (gynandromorph). Junior synonym of lobicornis: Dalla Torre, 1893: 111. Subspecies of scabrinodis: Emery, 1908a: 176; Bondroit, 1910: 495; Emery, 1916b: 120; Menozzi, 1922b: 325; Finzi, 1924a: 12; Finzi, 1926: 101; Menozzi, 1936d: 270. Status as species: Bondroit, 1918: 102; Santschi, 1931b: 345; Karavaiev, 1934: 80; Stitz, 1939: 94; Weber, 1939b: 144; Holgersen, 1940: 184; Novak & Sadil, 1941: 79; Bernard, 1967: 117; Baroni Urbani, 1971c: 31; Kutter, 1977c: 68; Arnol'di & Dlussky, 1978: 534; Collingwood, 1979: 54; Seifert, 1988b: 31; Atanassov & Dlussky, 1992: 97. Senior synonym of scabrinodolobicornis: Radchenko, 1994e: 80. See also: Emery, 1895c: 314; Radchenko & Elmes, 2010: 250.
- scabrinodolobicornis. Myrmica rubra var. scabrinodolobicornis Forel, 1874: 77 (w.q.m.) SWITZERLAND. Subspecies of scabrinodis: Emery, 1921f: 40; of sabuleti: Weber, 1948a: 296. Junior synonym of lonae: Sadil, 1952: 249; of sabuleti: Radchenko, 1994e: 80.
Taxonomic Notes
Ebsen et al (2019) - Myrmica lonae was recently resurrected as a species separate from M. sabuleti (Seifert, 2000), but where our mtDNA data could not find any evidence to support this. No specific M. lonae haplotypes were found in the four sequenced specimens (Figs. 1 and 2), which suggests that they share the same gene pool as M. sabuleti, as most of the samples were from sympatric populations in The Netherlands and Hungary, and the remaining GenBank sequence came from Finland where only M. lonae has been recorded (Seifert, 2000). The similarity between M. lonae and M. sabuleti is remarkable, because all specimens shared the most abundant Cyt-B M. sabuleti haplotype (Fig. 3), whereas established closely related species (Myrmica hirsuta, Myrmica spinosior, Myrmica vandeli) were differentiated with good bootstrap support. The M. lonae phenotype is associated with colder and wetter habitats than typical M. sabuleti (Elmes et al., 1998; Seifert, 2000; Radchenko & Elmes, 2010; JRE, personal observation), suggesting that this may be an ecotype of M. sabuleti. This interpretation is also supported by the fact that M. lonae and M. sabuleti have identical cuticular hydrocarbon profiles, while other Myrmica species examined can be readily separated based on surface chemistry (Guillem, 2014). Given this overall pattern within the M. sabuleti complex, we hypothesise that the Spanish specimen of M. sabuleti may represent yet another cryptic species (the long branch towards the right in Fig. 2).
Unless otherwise noted the text for the remainder of this section is reported from the publication that includes the original description.
Description
Karyotype
- See additional details at the Ant Chromosome Database.
- Explore: Show all Karyotype data or Search these data. See also a list of all data tables or learn how data is managed.
- 2n = 46 (Switzerland) (Hauschteck, 1965).
- n = 23 (Switzerland) (Hauschteck-Jungen & Jungen, 1983).
Etymology
Radchenko and Elmes (2010) - name derived from the Latin word sabulum = sand, sandy soil, to indicate its habitat.
References
- Arnan, X., Cerdá, X., Retana, J. 2012. Distinctive life traits and distribution along environmental gradients of dominant and subordinate Mediterranean ant species. Oecologia 170, 489–500 (doi:10.1007/s00442-012-2315-y).
- Arnol'di, K. V.; Dlussky, G. M. 1978. Superfam. Formicoidea. 1. Fam. Formicidae - ants. Pp. 519-556 in: Medvedev, G. S. (ed.) Keys to the insects of the European part of the USSR. Vol. 3. Hymenoptera. Part 1. Opredeliteli Faune SSSR 119:3-584. (page 534, Status as species)
- Atanassov, N.; Dlussky, G. M. 1992. Fauna of Bulgaria. Hymenoptera, Formicidae. Fauna Bûlg. 22: 1-310 (page 97, Status as species)
- Barbero, F., Bonelli, S., Thomas, J.A., Balletto, E., Schonrogge, K. 2009. Acoustical mimicry in a predatory social parasite of ants. Journal of Experimental Biology 212, 4084–4090. (doi:10.1242/JEB.032912).
- Barbero, F., Patricelli, D., Witek, M., Balletto, E., Casacci, L.P., Sala, M., Bonelli, S. 2012. Myrmica Ants and Their Butterfly Parasites with Special Focus on the Acoustic Communication. Psyche: A Journal of Entomology 2012, 1–11 (doi:10.1155/2012/725237).
- Baroni Urbani, C. 1971c. Catalogo delle specie di Formicidae d'Italia (Studi sulla mirmecofauna d'Italia X). Memorie della Societa Entomologica Italiana 50: 5-287 (page 31, Status as species)
- Báthori, F., Rádai, Z., Tartally, A. 2017. The effect of Rickia wasmanniii (Ascomycota, Laboulbeniales) on the aggression and boldness of Myrmica scabrinodis (Hymenoptera, Formicidae). Journal of Hymenoptera Research. 58:41–52. (doi:10.3897/jhr.58.13253)
- Bátori, Z., Vojtkó, A., Maák, I.E., Lőrinczi, G., Farkas, T., Kántor, N., Tanács, E., Kiss, P.J., Juhász, O., Módra, G., Tölgyesi, C., Erdős, L., Aguilon, D.J., Keppel, G. 2019. Karst dolines provide diverse microhabitats for different functional groups in multiple phyla. Scientific Reports 9, 7176 (doi:10.1038/s41598-019-43603-x).
- Beckers R., Goss, S., Deneubourg, J.L., Pasteels, J.M. 1989. Colony size, communication and ant foraging Strategy. Psyche 96: 239-256 (doi:10.1155/1989/94279).
- Bereczki, J., Póliska, S., Váradi, A., Tóth, J.P. 2019. Incipient sympatric speciation via host race formation in Phengaris arion (Lepidoptera: Lycaenidae). Organisms Diversity, Evolution 20, 63–76 (doi:10.1007/s13127-019-00418-y).
- Bernadou, A., Fourcassié, V., Espadaler, X. 2013. A preliminary checklist of the ants (Hymenoptera, Formicidae) of Andorra. ZooKeys 277, 13–23 (doi:10.3897/zookeys.277.4684).
- Bernard, F. 1967a [1968]. Faune de l'Europe et du Bassin Méditerranéen. 3. Les fourmis (Hymenoptera Formicidae) d'Europe occidentale et septentrionale. Paris: Masson, 411 pp. (page 117, Status as species, Senior synonym of lonae)
- Blatrix, R., Aubert, C., Decaëns, T., Berquier, C., Andrei-Ruiz, M.-C., Galkowski, C. 2020. Contribution of a DNA barcode to an assessment of the specificity of ant taxa (Hymenoptera: Formicidae) on Corsica. European Journal of Entomology 117: 420-429 (doi:10.14411/eje.2020.046).
- Boer, P. 2008. Observations of summit disease in Formica rufa Linnaeus, 1761 (Hymenoptera: Formicidae). Myrmecological News 11. 63-66.
- Bolton, B. 1995b. A new general catalogue of the ants of the world. Cambridge, Mass.: Harvard University Press, 504 pp. (page 282, Senior synonym of scabrinodolobicornis: new synonymy)
- Bondroit, J. 1910 [1909]. Les fourmis de Belgique. Ann. Soc. Entomol. Belg. 53: 479-500 (page 495, Variety/subspecies of scabrinodis)
- Bondroit, J. 1918. Les fourmis de France et de Belgique. Ann. Soc. Entomol. Fr. 87: 1-174 (page 102, Status as species)
- Borowiec, L. 2014. Catalogue of ants of Europe, the Mediterranean Basin and adjacent regions (Hymenoptera: Formicidae). Genus (Wroclaw) 25(1-2): 1-340.
- Borowiec, L., Lebas, C., Salata, S. 2022. Notes on ants (Hymenoptera: Formicidae) from three northern Aegean islands – Lemnos, Samothraki and Thasos. Annals of the Upper Silesian Museum in Bytom, Entomology 31: 1-14 (doi:10.5281/ZENODO.7346453).
- Borowiec, L., Salata, S. 2017. First certain record of Lasius alienus (F RSTER, 1850) (Hymenoptera: Formicidae) and three ant species new to the Republic of Macedonia. Acta Entomologica Silesiana 25(online 019): 1–4.
- Cantone S. 2017. Winged Ants, The Male, Dichotomous key to genera of winged male ants in the World, Behavioral ecology of mating flight (self-published).
- Carroll, T.M. 2011. The ants of Indiana (Hymenoptera: Formicidae). M.S. thesis, Purdue University.
- Casevitz-Weulersse, J. 1990b. Étude systématique de la myrmécofaune corse (Hymenoptera, Formicidae) (Première partie). Bull. Mus. Natl. Hist. Nat. Sect. A Zool. Biol. Écol. Anim. (4) 12: 135-163 (page 137, Senior synonym of spinosior)
- Castracani, C., Spotti, F.A., Schifani, E., Giannetti, D., Ghizzoni, M., Grasso, D.A., Mori, A. 2020. Public engagement provides first insights on Po Plain ant communities and reveals the ubiquity of the cryptic species Tetramorium immigrans (Hymenoptera, Formicidae). Insects 11, 678. (doi:10.3390/insects11100678).
- Collingwood, C. A. 1979. The Formicidae (Hymenoptera) of Fennoscandia and Denmark. Fauna Entomol. Scand. 8: 1-174 (page 54, Status as species)
- Collingwood, C.A., Prince, A. 1998. A guide to ants of Continental Portugal (Hymenoptera: Formicidae). Boletim da Sociedade Portuguesa de Entomologia. Supl nº5, pp 49.
- Csata, E., Dussutour, A. 2019. Nutrient regulation in ants (Hymenoptera: Formicidae): a review. Myrmecological News 29: 111-124 (doi:10.25849/MYRMECOL.NEWS_029:111).
- Csősz, S., Báthori, F., Gallé, L., Lőrinczi, G., Maák, I., Tartally, A., Kovács, É., Somogyi, A.Á., Markó, B. 2021. The myrmecofauna (Hymenoptera: Formicidae) of Hungary: Survey of ant species with an annotated synonymic inventory. Insects 16;12(1):78 (doi:10.3390/insects12010078).
- Csosz, S., Marko, B., Galle, L. 2011. The myrmecofauna (Hymenoptera: Formicidae) of Hungary: an updated checklist. North-Western Journal of Zoology 7: 55-62.
- Czechowska, W., Radchenko, A. 1997. Myrmica hirsuta Elmes, 1978 (Hymenoptera, Formicidae) - a socially parasitic ant species new to Poland. Fragmenta Faunistica 40: 53-57.
- Czechowski, W., Czechowska, W., Radchenko, A. 2007. Strikingly malformed host morphology: Myrmica rugulosa Nyl. and Myrmica sabuleti Mein. (Hymenoptera: Formicidae) parasitised by mermithid nematodes. Fragmenta Faunistica 50: 139–148.
- Czechowski, W., Radchenko, A., Czechowska, W. 2002. The ants (Hymenoptera, Formicidae) of Poland. MIZ PAS Warsaw.
- Czekes, Z., Radchenko, A.G., Csosz, S., sz-Len, A.S., Tausan, I., Benedek, K., Mark, B. 2012. The ant genus Myrmica Latreille, 1804 (Hymenoptera: Formicidae) in Romania: distribution of species and key for their identification. Entomologica Romanica 17: 29-50.
- Dalla Torre, K. W. von. 1893. Catalogus Hymenopterorum hucusque descriptorum systematicus et synonymicus. Vol. 7. Formicidae (Heterogyna). Leipzig: W. Engelmann, 289 pp. (page 111, Junior synonym of lobicornis)
- Dekoninck, W., Ignace, D., Vankerkhoven, F., Wegnez, P. 2012. Verspreidingsatlas van de mieren van België. Bulletin de la Société royale belge d’Entomologie 148: 95-186.
- Donisthorpe, H. 1915d. Descriptions of a pterergate and two gynandromorphs of Myrmica scabrinodis Nyl., with a list of all the known cases of the latter. Entomol. Rec. J. Var. 27: 258-260 (page 258, gynandromorph described)
- Ebsen, J.R., Boomsma, J.J. & Nash, D.R. 2019. Phylogeography and cryptic speciation in the Myrmica scabrinodis Nylander, 1846 species complex (Hymenoptera: Formicidae), and their conservation implications. Insect Conservation and Diversity 12: 467-480 (doi:10.1111/icad.12366).
- Emery, C. 1908a. Beiträge zur Monographie der Formiciden des paläarktischen Faunengebietes. Dtsch. Entomol. Z. 1908: 165-205 (page 176, queen described, Variety/subspecies of scabrinodis; page 177, Senior synonym of granulinodis (footnote))
- Emery, C. 1916a [1915]. Fauna entomologica italiana. I. Hymenoptera.-Formicidae. Bull. Soc. Entomol. Ital. 47: 79-275 (page 120, Variety/subspecies of scabrinodis)
- Espadaler, X., Santamaria, S. 2012. Ecto- and Endoparasitic Fungi on Ants from the Holarctic Region. Psyche Article ID 168478, 10 pages (doi:10.1155/2012/168478).
- Finzi, B. 1924a. Formiche dell'isola d'Elba e Monte Argentario. Boll. Soc. Entomol. Ital. 56: 12-15 (page 12, Variety/subspecies of scabrinodis)
- Finzi, B. 1926. Le forme europee del genere Myrmica Latr. Primo contributo. Boll. Soc. Adriat. Sci. Nat. Trieste 29: 71-119 (page 101, Variety/subspecies of scabrinodis)
- García, F., Arnal, J.M., Espadaler, X. 2008. Primeros registros de Myrmica bibikoffi Kutter, 1963 (Hymenoptera: Formicidae) en la Península Ibérica. Heteropterus Rev. Entomol. 8, 211–215.
- García, F., Cuesta-Segura, A.D., Espadaler, X. 2024. Myrmica babiensis sp. nov. (Hymenoptera: Formicidae), a new social parasite from the NW Iberian Peninsula. Annales Zoologici 74(1), 113-127 (doi:10.3161/00034541anz2024.74.1.006).
- Giannetti, D., Schifani, E., Castracani, C., Ghizzoni, M., Delaiti, M., Pfenner, F., Spotti, F.A., Mori, A., Ioriatti, C., Grasso, D.A. 2021. Assessing ant diversity in agroecosystems: the case of Italian vineyards of the Adige Valley. Redia 104, 97–109 (doi:10.19263/redia-104.21.11).
- Glaser, F. 2016. Artenspektrum, Habitatbindung und naturschutzfachliche Bedeutung von Ameisen (Hymenoptera, Formicidae) am Stutzberg (Vorarlberg, Österreich). inatura – Forschung 34: 26 S.
- Haelewaters, D., Boer, P., Noordijk, J. 2015. Studies of Laboulbeniales (Fungi, Ascomycota) on Myrmica ants: Rickia wasmanniii in the Netherlands. Journal of Hymenoptera Research 44, 39–47 (doi:10.3897/jhr.44.4951).
- Hauschteck, E. 1965. Halbe haploide Chromosomenzahl im Hoden von Myrmica sulcinodis Nyl. (Formicidae). Experientia (Basel) 21: 323-325 (page 325, karyotype described)
- Holgersen, H. 1940. Myrmekologiske notiser I. Nor. Entomol. Tidsskr. 5: 183-187 (page 184, Status as species)
- Jansen, G., Savolainen, R. 2010. Molecular phylogeny of the ant tribe Myrmicini (Hymenoptera: Formicidae). Zoological Journal of the Linnean Society 160(3), 482–495 (doi:10.1111/j.1096-3642.2009.00604.x).
- Jansen, G., Savolainen, R., Vepsäläinen, K. 2010. Phylogeny, divergence-time estimation, biogeography and social parasite–host relationships of the Holarctic ant genus Myrmica (Hymenoptera: Formicidae). Molecular Phylogenetics and Evolution 561, 294–304 (doi:10.1016/j.ympev.2010.01.029).
- Kanizsai, O. 2014. Forms and background factors of the coexistence between colonies of Formica fusca and Camponotus vagus (Hymenoptera, Formicidae) in forest steppe habitats. Ph.D. thesis, University of Szeged.
- Karaman, C., Kiran, K. 2022. Additional records of parasitic Camponotus Mayr (Hymenoptera: Formicidae) species from Turkey with queen description of Camponotus ruseni Karaman, 2012. Zoology in the Middle East 68(2), 156–164 (doi:10.1080/09397140.2022.2051918).
- Karavaiev, V. 1934. The fauna of the family Formicidae (ants) of the Ukraine. Tr. Inst. Zool. Biol. Vseukr. Akad. Nauk Ser. 1 Pr. Syst. Faun. 1934: 1-164 (page 80, Status as species)
- Kiran, K., Karaman, C. 2020. Additions to the ant fauna of Turkey (Hymenoptera, Formicidae). Zoosystema 42(18), 285-329 (doi:10.5252/zoosystema2020v42a18).
- Kirchmair, G., Friess, T. et al. 2017. Zoologischer Bericht vom Tag der Biodiversität 2017 im Naturpark Südsteiermark. Mitteilungen des Naturwissenschaftlichen Vereines für Steiermark 147: 99–134.
- Kutter, H. 1977c. Hymenoptera, Formicidae. Insecta Helv. Fauna 6: 1-298 (page 68, Status as species)
- Laciny, A. 2021. Among the shapeshifters: parasite-induced morphologies in ants (Hymenoptera, Formicidae) and their relevance within the EcoEvoDevo framework. EvoDevo 12, 2 (doi:10.1186/s13227-021-00173-2).
- Lapeva-Gjonova, A., Antonova, V. 2022. An updated checklist of ants (Hymenoptera, Formicidae) of Bulgaria, after 130 years of research. Biodiversity Data Journal 10, e95599 (doi:10.3897/bdj.10.e95599).
- Lapeva-Gjonova, A., Antonova, V., Ljubomirov, T. 2021. Ants (Hymenoptera, Formicidae) of Sarnena Sredna Gora Mountains (Bulgaria). Fauna of Sarnena Sredna Gora Mts, Part 2 ZooNotes, Supplement 10: 18-27.
- Lapeva-Gjonova, A., Kiran, K. 2012. Ant fauna (Hymenoptera, Formicidae) of Strandzha (Istranca) Mountain and adjacent Black Sea coast. North-Western Journal of Zoology 8(1), 72-84.
- Maziarz, M., Broughton, R.K., Casacci, L.P., Dubiec, A., Maák, I., Witek, M. 2020. Thermal ecosystem engineering by songbirds promotes a symbiotic relationship with ants. Scientific Reports 10, 20330 (doi:10.1038/s41598-020-77360-z).
- Meinert, F. 1861. Bidrag til de danske Myrers Naturhistorie. K. Dan. Vidensk. Selsk. Skr. (5) 5: 273-340 (page 327, worker, male described)
- Menozzi, C. 1922c. Contribution à la faune myrmécologique de l'Espagne. Bol. R. Soc. Esp. Hist. Nat. 22: 324-332 (page 325, Variety/subspecies of scabrinodis)
- Menozzi, C. 1936b. Nuovi contributi alla conoscenza della fauna delle Isole italiane dell'Egeo. VI. Hymenoptera - Formicidae. Boll. Lab. Zool. Gen. Agrar. R. Sc. Super. Agric. 29: 262-311 (page 270, Variety/subspecies of scabrinodis)
- Nemet, E., Czekes, Z., Tausan, I., Marko, B. 2012. Contribution to the knowledge of the myrmecofauna of the Cefa Nature Park (North-Western Romania). Acta Scientiarum Transylvanica Biologia 20, 61-72.
- Novák, V.; Sadil, J. 1941. Klíc k urcování mravencu strední Evropy se zvlástním zretelem k mravencí zvírene Cech a Moravy. Entomol. Listy 4: 65-115 (page 79, Status as species)
- Obregon, R., M. R. Shaw, J. Fernandez-Haeger, and D. Jordano. 2015. Parasitoid and ant interactions of some Iberian butterflies (Insecta: Lepidoptera). Shilap-Revista De Lepidopterologia. 43:439-454.
- Purkart, A., Kollár, J., Goffová, K. 2019. Fauna of ants (Hymenoptera: Formicidae) of selected sand habitats in Podunajsko Region, Slovakia. Naturae Tutela 23(1): 101-111.
- Radchenko, A. G. 1994h. Survey of the species of the rubra, rugosa, arnoldii, luteola and schencki groups of the genus Myrmica (Hymenoptera, Formicidae) from central and eastern Palearctic. Zool. Zh. 73(1 11: 72-80 (page 80, senior synonym of scabrinodolobicornis and scabrinodosaubuleti)
- Radchenko, A.G. & Elmes, G.W. 2010. Myrmica ants of the Old World. Fauna Mundi 3: 1-789.
- Rericha, L. 2007. Ants of Indiana. Indiana Department of Natural Resources, 51pp.
- Sadil, J. V. 1952 [1951]. A revision of the Czechoslovak forms of the genus Myrmica Latr. (Hym.). Sb. Entomol. Oddel. Nár. Mus. Praze 27: 233-278 (page 246, if synonymy correct then granulinodis has priority))
- Santschi, F. 1931c. Notes sur le genre Myrmica (Latreille). Rev. Suisse Zool. 38: 335-355 (page 345, Status as species)
- Savolainen, R., Vepsalainen, K. 2003. Sympatric speciation through intraspecific social parasitism. Proceedings of the National Academy of Sciences 100(12): 7169–7174.
- Schifani, E., Csősz, S., Viviano, R., Alicata, A. 2021. Ant diversity on the largest Mediterranean islands: on the presence or absence of 28 species in Sicily (Hymenoptera, Formicidae). Natural History Sciences 8, 55–70 (doi:10.4081/nhs.2021.532).
- Schifani, E., Nalini, E., Gentile, V., Alamanni, F., Ancona, C., Caria, M., Cillo, D., Bazzato, E. 2021. Ants of Sardinia: An updated checklist based on new faunistic, morphological and biogeographical notes. Redia 104, 21–35 (doi:10.19263/redia-104.21.03).
- Schultner, E., Pulliainen, U. 2020. Brood recognition and discrimination in ants. Insectes Sociaux 67, 11–34 (doi:10.1007/s00040-019-00747-3).
- Seifert, B. 1988b. A taxonomic revision of the Myrmica species of Europe, Asia Minor, and Caucasia (Hymenoptera, Formicidae). Abh. Ber. Naturkundemus. Görlitz 62(3): 1-75 (page 31, Status as species, Senior synonym of lonae, spinosior)
- Steiner, F.M., Kohler, G., Seifert, B., Arthofer, W., Schlick-Steiner, B.C., Buschinger, A. 2019. A worker-like female of Myrmica sabuleti Meinert, 1861 (Hymenoptera: Formicidae: Myrmicinae) in a pitfall trap with five mermithids (Nematoda: Mermithidae) protruding from the gaster. Sociobiology 66(3): 400-407 (doi:10.13102/sociobiology.v66i3.4338).
- Steiner, F.M., Schlick-Steiner, B.C., Holzinger, W., Komposch, C., Pazoutova, S., Sanetra, M., Christian, E. 2004. A novel relationship between ants and a leafhopper (Hymenoptera: Formicidae; Hemiptera: Cicadellidae). European Journal of Entomology 101, 689-692.
- Stevens, I., Vankerkhoven, F., Vanormeligen, P., Dekoninck, W. 2020. New observations of the social parasitic ant Myrmica karavajevi (Arnoldi, 1930) (Hymenoptera: Formicidae) in Belgium. Bulletin de la Société royale belge d’Entomologie 156: 113–121
- Stitz, H. 1939. Die Tierwelt Deutschlands und der angrenzenden Meersteile nach ihren Merkmalen und nach ihrer Lebensweise. 37. Theil. Hautflüger oder Hymenoptera. I: Ameisen oder Formicidae. Jena: G. Fischer, 428 pp. (page 94, Status as species)
- Tartally, A., Somogyi, A.Á., Révész, T., Nash, D.R. 2020. Host ant change of a socially parasitic butterfly (Phengaris alcon) through host nest take-over. Insects 11, 556 (doi:10.3390/INSECTS11090556).
- Weber, N. A. 1939b. Description of new North American species and subspecies of Myrmica Latreille (Hym.: Formicidae). Lloydia 2: 144-152 (page 144, Status as species)
- Wegnez, P. 2017. Découverte de Myrmica lobicornis Nylander, 1846 et Lasius jensi Seifert, 1982, deux nouvelles espèces pour le Grand-Duché de Luxembourg (Hymenoptera: Formicidae). Bulletin de la Société royale belge d’Entomologie153, 46–49.
- Wehner, R. 2009. The architecture of the desert ant's navigational toolkit (Hymenoptera: Formicidae). Myrmecological News 12, 85-96.
- Wiezik, M., Svitok, M., Wieziková, A., Dovčiak, M. 2013. Shrub encroachment alters composition and diversity of ant communities in abandoned grasslands of western Carpathians. Biodiversity and Conservation 22, 2305–2320 (doi:10.1007/s10531-013-0446-z).
References based on Global Ant Biodiversity Informatics
- Agosti D. 1983. Zur Insektenfauna der Umgebung der Vogelwarte Sempach, Kanton Luzern. XIII. Hymenoptera 2: Formicidae (Ameisen). Entomologische Berichte Luzern 10: 91-92.
- Agosti, D. and C.A. Collingwood. 1987. A provisional list of the Balkan ants (Hym. Formicidae) and a key to the worker caste. I. Synonymic list. Mitteilungen der Schweizerischen Entomologischen Gesellschaft, 60: 51-62
- Allen G. W. 1989. A key to the worker castes of the ants of Kent (Hymenoptera: Formicidae). Transactions of the Kent Field Club 11: 8-23.
- Alvarado M., and L. Galle. 2000. Ant assemblages associated with lowland forests in the southern part of the great Hungarian plain. Acta Zoologica Academiae Scientarum Hungaricae 46(2): 79-102.
- Andoni V. 1977. Kontribut mbi Himenopteret e familjes Formicidae te vendit tone. Buletini I Shkencave te Natyres 31(2): 93-101.
- AntArea. Accessed on February 5th 2014 at http://antarea.fr/fourmi/
- Antarea (at www.antarea.fr on June 11th 2017)
- Antonova V., and L. Penev. 2008. Classification of assemblages of ants in the green areas in Sofia City. Acta Zoologica Bulgarica 60(2): 103-110.
- ArtDatabanken Bugs (via GBIG)
- Asociacion Iberica de Mirmecologia. 2011. List of species collected during the Taxomara Lisboa 2011. Iberomyrmex 3: 30-31.
- Asociacion Iberica de Mirmecologia. 2014. List of species collected during the Taxomara 2014 Oviedo. Iberomyrmex 6: 23-24.
- Assing V. 1989. Die Ameisenfauna (Hym.: Formicidae) nordwestdeutscher Calluna-Heiden. Drosera 89: 49-62.
- Bare O. S. 1929. A taxonomic study of Nebraska ants, or Formicidae (Hymenoptera). Thesis, University of Nebraska, Lincoln, USA.
- Baroni Urbani C., and C. A. Collingwood. 1976. A Numerical Analysis of the Distribution of British Formicidae (Hymenoptera, Aculeata). Verhandlungen der Naturforschenden Gesellschaft in Basel 85: 51-91.
- Baroni Urbani C., and C. A. Collingwood. 1977. The zoogeography of ants (Hymenoptera, Formicidae) in Northern Europe. Acta Zoologica Fennica 152: 1-34.
- Baroni Urbani, C. "Studi sulla mirmecofauna d'Italia. II. Formiche di Sicilia." Atti dell'Accademia Gioenia di Scienze Naturali in Catania (6) 16 (1964): 25-66.
- Barrett K. E. 1967. Ants in South Brittany. Entomologist's Record and Journal of Variation 79:112-116.
- Barrett K. E. J. 1968. Ants in western France. Entomologist 101: 153-155.
- Barrett K. E. J. 1968b. The distribution of ants in central southern England. Transactions of the Society for British Entomology 17: 235-250.
- Barrett K. E. J. 1970. Ants in France, 1968-69. Entomologist 103: 270-274.
- Baugnee J. Y. 2003. Camponotus piceus (Leach, 1825), fourmi nouvelle pour la faune belge decouverte dans le parc naturel Viroin-Hermeton (Hymenoptera: Formicidae). Bulletin S. R. B. E./K. B. V. E. 139: 219-225.
- Behr D., S. Lippke, and K. Colln. 1996. Zur kenntnis der ameisen von Koln (Hymenoptera, Formicidae). Decheniana-Beihefte (Bonn) 35: 215-232.
- Behr D., and K. Colln. 1993. Zur ameisenfauna (Hymenoptera, Formicidae) von Gonnersdorf (Kr. Daun). Dendrocopos 20: 148-160.
- Bernadou A., V. Fourcassié, and X. Espadaler. 2013. A preliminary checklist of the ants (Hymenoptera, Formicidae) of Andorra. Zookeys 277: 13-23.
- Bernadou A., X. Espadaler, A. Le Goff, and V. Fourcassie. 2015. Ant community organization along elevational gradients in a temperate ecosystem. Insect. Soc. 62:5971
- Bernadou, A., G. Latil, V. Fourcassié, and X. Espadaler. "Les formigues de la Vall del Madriu-Perafita-Claror : diversitat i distribució." Hàbitats, 13 (2006): 10-21.
- Bernard F. 1960. Fourmis récoltées en Corse par J. Bonfils (1957). Compte Rendu Sommaire des Séances de la Société de Biogéographie 36: 108-114.
- Bernard F. 1967. Faune de l'Europe et du Bassin Méditerranéen. 3. Les fourmis (Hymenoptera Formicidae) d'Europe occidentale et septentrionale. Paris: Masson, 411 pp.
- Berville L., C. Santelli, J. Reybaud, M. Renucci, P. Ponel, O. Blight, and E. Provost. 2014. Suivi d’un site atelier dans le golfe de Fos: Une diversite myrmecologique insoupconne. Etudes Vauclusiennes 82: 71-78.
- Bezdecka P. 1996. The ants of Slovakia (Hymenoptera: Formicidae). Entomofauna carpathica 8: 108-114.
- Bezdeckova K., and P. Bezdecka. 2009. Nejvetsi polykalicka kolonie Formica foreli (Hymenoptera: Formicidae) y Ceské republice. Acta rerum naturalium 7: 121126.
- Bigot L. 1959. Complement a l'inventaire de la faune entomologique de la Camargue 93eme note). La Terre et la Vie 106 : 149-157.
- Blacker N. C. 1989. The ants (Hymenoptera, Formicidae) of the Gower Peninsula, West Glamorgan, South Wales. Entomologist's Record and Journal of Variation 101: 261-266.
- Blacker N. C. and C. A. Collingwood. 2002. Some significant new records of ants (Hymenoptera: Formicidae) from the Salisbury area, south Wiltshire, England, with a key to the British species of Lasius. British Journal of Entomology and Natural History 15: 25-46
- Blatrix R., C. Lebas, C. Galkowski, P. Wegnez, P. Pimenta, and D. Morichon. 2016. Vegetation cover and elevation drive diversity and composition of ant communities (Hymenoptera: Formicidae) in a Mediterranean ecosystem. – Myrmecological News 22: 119-127.
- Boer P. 2019. Species list of the Netherlands. Accessed on January 22 2019 at http://www.nlmieren.nl/websitepages/specieslist.html
- Boer P., W. Dekoninck, A. J. Van Loon, and F. Vankerkhoven. 2003. Lijst van mieren (Hymenoptera: Formicidae) van Belgie en Nederland, hun Nederlandse namen en hun voorkomen. Entomologische Berichten (Amsterdam) 63: 54-58.
- Boer P., W. Dekoninck, A. J. van Loon, and F. Vankerkhoven. 2003. Lijst van mieren (Hymenoptera: Formicidae) van Belgie en Nederland, hun Nederlandse namen en hun voorkomen. Entomologische Berichten 63(3): 54-57.
- Boer P., W. Dekoninck, A. J. van Loon, and F. Vankerkhoven. 2003. List of ants (Hymenoptera: Formicidae) of Belgium and The Netherlands, their status and Dutch vernacular names. Entomologische Berichten 63 (3): 54-58.
- Boer P., and J. Noordijk. 2004. De ruige gaststeekmier Myrmica hirsuta nieuw voor Nederland (Hymenoptera: Formicidae). Ned. Faun. Meded. 20: 25-32.
- Boer P., and J. Noordijk. 2005. Myrmica schenckioides nov. sp., a new socially parasitic ant species (Hymenoptera: Formicidae). Entomol. Ber. (Amst.) 65(4): 120-123.
- Bonaric J. C. 1971. Contribution a l'etude systematique et ecologique des formicides du Bas-Languedoc. PhD thesis Universite des sciences et techniques du Languedoc, 175 pages.
- Bonaric J. C. 1971. Étude systématique et écologique des fourmis de lHérault. Ann. Soc. Hortic. Hist. Nat. Hérault 111: 81-87.
- Bonte D., W. Dekoninck, S. Provoost, E. Cosijns, and M. Hoffmann. 2003. Microgeographical distribution of ants (Hymenoptera: Formicidae) in coastal dune grassland and their relation to the soil structure and vegetation. Animal Biology 53(4): 367-377.
- Borowiec L. 2014. Catalogue of ants of Europe, the Mediterranean Basin and adjacent regions (Hymenoptera: Formicidae). Genus (Wroclaw) 25(1-2): 1-340.
- Borowiec L., and S. Salata. 2012. Ants of Greece - Checklist, comments and new faunistic data (Hymenoptera: Formicidae). Genus 23(4): 461-563.
- Bracko G. 2007. Checklist of the ants of Slovenia (Hymenoptera: Formicidae). Natura Sloveniae 9: 15-24
- Bracko G., K. Kiran, C. Karaman, S. Salata, and L. Borowiec. 2016. Survey of the ants (Hymenoptera: Formicidae) of the Greek Thrace. Biodiversity Data Journal 4: e7945. doi: 10.3897/BDJ.4.e7945
- Bracko, G. 2006. Review of the ant fauna (Hymenoptera:Formicidae) of Croatia. Acta Entomologica Slovenica 14(2): 131-156.
- Bracko, G. "Review of the ant fauna (Hymenoptera: Formicidae) of Croatia." Acta Entomologica Slovenica Vol 14 st (2006): 131-156.
- Braschler, B. and B. Baur. 2005. Experimental Small-Scale Grassland Fragmentation Alters Competitive Interactions among Ant Species. Oecologia 143(2):291-300
- Cammell, M. E., M. J. Way, and M. R. Paiva. "Diversity and structure of ant communities associated with oak, pine, eucalyptus and arable habitats in Portugal." Insectes Sociaux 43 (1996): 37-46.
- Carniel A. 1998. Ricerche sulla mirmecofauna delle Prealpi Orobiche (Lombardia) (Insecta, Hymenoptera, Formicidae). Atti. Mus. Civ. Stor. Nat. Morbegno 9: 29-39.
- Casevitz-Weulersse J. 1990. Etude Systematique de la Myrmecofaune Corse (Hymenoptera: Formicidae), deuxieme partie. Bull. Mus. Natn. Hist. Nat. Paris. 4eme serie 12, section A(2): 415-442.
- Casevitz-Weulersse J., and C. Galkowski. 2009. Liste actualisee des Fourmis de France (Hymenoptera, Formicidae). Bull. Soc. Entomol. Fr. 114: 475-510.
- Casevitz-Weulersse J., and M. Prost. 1991. Fourmis de la Côte-d'Or présentes dans les collections du Muséum d'Histoire Naturelle de Dijon. Bulletin Scientifique de Bourgogne 44: 53-72.
- Casevitz-Weulersse, J. "Contribution a la connaisance des fourmis de la Corse (Hymenoptera: Formicidae)." These de Doctorat Museum Nat (1989): 379pp.
- Castillo-Miralbes M. 2001. Artropodos presentes en carrona de cerdos en la comarca de la litera (Huesca). Bol. S.E.A. 28: 133-140.
- Cherix D., and S. Higashi. 1979. Distribution verticale des fourmis dans le Jura vaudois et recensement prelimaire des bourdons (Hymenoptera, Formicidae et Apidae). Bull. Soc. Vaud. Sc. Nat. 356(74): 315-324.
- Chevrier M., and C. Mouquet C. 2005. Etude des peuplements des invertébrés des dunes de Bretagne. Rapport GRETIA, avenant au Contrat-Nature 2, Conseil Régional de Bretagne, Conseils Généraux des Côtes d'Armor, du Finistère et du Morbihan : 127 p.
- Colindre L. 2015. Les fourmis en Picardie: bilan 2014 (Hymenoptera/ Formicidae). Entomologiste Picard 26, 15 pages.
- Colindre L. 2017. Richess et utilite du cortege de fourmis en foret d'Ermenonville, Oise, Region Hauts-de-France. Association des Entomologistes de Picardie. 19 pages.
- Collingwood C. A. 1951. The distribution of ants in north-west Scotland. Scottish Naturalist 63: 45-49
- Collingwood C. A. 1951. The distribution of ants in north-west Scotland. Scottish Naturalist 63: 45-49.
- Collingwood C. A. 1955. Ants in S.W. Scotland. Entomologist's Record and Journal of Variation 67: 11-12.
- Collingwood C. A. 1956. Ant hunting in France. Entomologist 89: 106-108.
- Collingwood C. A. 1961. Ants in the Scottish Highlands. Scotish Naturalist 70: 12-21.
- Collingwood C. A. 1971. A synopsis of the Formicidae of north Europe. Entomologist 104: 150-176
- Collingwood C. A., and J. E. Satchell. 1956. The ants of the South Lake District. Journal of the Society for British Entomology 5: 159-164.
- Collingwood C. A., and J. Hughes. 1987. Ant species in Yorkshire, England. Naturalist (Leeds) 112: 95-101.
- Collingwood C., and A. Prince. 1998. A guide to ants of continental Portugal (Hymenoptera: Formicidae). Boletim da Sociedade Portuguesa de Entomologia. Suplemento 5: 1-49.
- Collingwood C.A. 1955. Ants in S.W. Scotland. Entomol.Rec. 67: 11-12
- Collingwood C.A. 1959. Ants in the Scottish Highlands. The Scottish Naturalist. 70: 12-21
- Collingwood C.A. 1959. Scandinavian Ants. Entomol. Rec. 71: 78-83
- Collingwood C.A. 1961. New Vice-County Records for British Ants. Entomologist. 73: 90-93
- Collingwood C.A. and Satchell J.E. 1956. The Ants of the South Lake District. Journal of the Society for British Entomology. 5: 159-164
- Collingwood, C. A. 1958. The ants of the genus Myrmica in Britain. Proc. R. Entomol. Soc. Lond. Ser. A 33: 65-75
- Collingwood, C. A. 1958b. A key to the species of ants (Hymenoptera, Formicidae) found in Britain. Trans. Soc. Br. Entomol. 13: 69-96
- Collingwood, C. A. 1964. The Identification of British Ants (Hym. Formicidae). Transactions of the Society for British Entomology. 16:93-121.
- Collingwood, C. A. 1974. A revised list of Norwegian ants (Hymenoptera: Formicidae). Norsk Entomologisk Tidsskrift 21: 31-35.
- Collingwood, C. A., and I. H. H. Yarrow. "A survey of Iberian Formicidae." EOS (Revista española de entomología) 44 (1969): 53-101.
- Collingwood, C. A. "The Formicidae (Hymenoptera) of Fennoscandia and Denmark." Fauna Entomologica Scandinavica 8 (1979): 1-174.
- Collingwood, C.A. 1958. A survey of Irish Formicidae. Proceedings of the Royal Irish Academy 59B:213-219
- Consani M., and P. Zangheri. 1952. Fauna di Romagna. Imenotteri - Formicidi. Memorie della Societa Entomologica Italiana 31: 38-48.
- Csosz S., B. Marko, K. Kiss, A. Tartally, and L. Galle. 2002. The ant fauna of the Ferto-Hansag National Park (Hymenoptera: Formicidae). In: Mahunka, S. (Ed.): The fauna of the Fert?-Hanság National Park. Hungarian Natural History Museum, Budapest, pp. 617-629.
- Csősz S., B. Markó, and L. Gallé. 2001. Ants (Hymenoptera: Formicidae) of Stana Valley (Romania): Evaluation of the effectiveness of a myrmecological survey. Entomologica Romanica 6 : 121-126.
- Csősz S., B. Markó, and L. Gallé. 2011. The myrmecofauna (Hymenoptera: Formicidae) of Hungary: an updated checklist. North-Western Journal of Zoology 7: 55-62.
- Czechowska W., and A. Radchenko. 1997. Myrmica hirsuta Elmes, 1978 (Hymenoptera, Formicidae) - a socially parasitic ant species new to Poland. Fragmenta Faunistica 40: 53-57.
- Czechowski W., A. Radchenko, W. Czechowska and K. Vepsäläinen. 2012. The ants of Poland with reference to the myrmecofauna of Europe. Fauna Poloniae 4. Warsaw: Natura Optima Dux Foundation, 1-496 pp
- Czekes Z., Radchenko, A. G., Csősz, S. Szász-Len, A., Tăuşan, I., Benedek, K., and Markó, B. 2013. The genus Myrmica Latreille, 1804 (Hymenoptera: Formicidae) in Romania: distribution of species and key for their identification. Entomologica Romanica 17: 29-50.
- Davis W. T., and J. Bequaert. 1922. An annoted list of the ants of Staten Island and Long Island, N. Y. Bulletin of the Brooklyn Entomological Society 17(1): 1-25.
- Dekoninck W., F. Hendrickx, M. Dethier, and J. P. Maelfait. 2010. Forest Succession Endangers the Special Ant Fauna of Abandoned Quarries along the River Meuse (Wallonia, Belgium). Restoration Ecology 18(5): 681690.
- Dekoninck W., H. De Koninck, J. Y. Baugnee, and J. P. Maelfait. 2007. Ant biodiversity conservation in Belgian calcareous grasslands: active management is vital. Belg. J. Zool. 137 (2): 137-146.
- Dekoninck W., K. Desender, and P. Grootaert. 2008. Establishment of ant communities in forests growing on former agricultural fields: Colonisation and 25 years of management are not enough (Hymenoptera: Formicidae). Eur. J. Entomol. 105: 681689.
- Dekoninck W., P. Boer, and J. P. Maelfait. 2004. Lasius platythorax Seifert, 1991 as a host of several Chthonolasius species, with remarks on the colony foundation of the parasites (Hymenoptera: Formicidae). Myrmecologische Nachrichten 6: 5-8.
- Della Santa E. 1994. Guide pour l'identification des principales espèces de fourmis de Suisse. Miscellanea Faunistica Helvetiae 3: 1-124.
- Della Santa E. 1995. Fourmis de Provence. Faune Provence 16: 5-37.
- Della Santa E. 2000. L'identification des espèces du genre Myrmica Latreille (Formicidae) de Suisse; essai de présentation synoptique. Bull. Romand Entomol. 18: 169-187.
- Dewes E. 2005. Ameisenerfassung im Waldschutzgebiet Steinbachtal/Netzbachtal. Abh. Delattinia 31: 89-118.
- Donisthorpe H. 1914. Myrmecophilous notes for 1913. Entomologist's Record and Journal of Variation 26: 37-45.
- Du Merle P. 1978. Les peuplements de fourmis et les peuplements d'acridiens du Mont Ventoux II. - Les peuplements de fourmis. Terre Vie 32(1): 161-218.
- Dubovikoff D. A., and Z. M. Yusupov. 2018. Family Formicidae - Ants. In Belokobylskij S. A. and A. S. Lelej: Annotated catalogue of the Hymenoptera of Russia. Proceedingss of the Zoological Institute of the Russian Academy of Sciences 6: 197-210.
- Dvorak, L., P. BOGUSCH, I. MALENOVSKÝ, P. BEZDÌÈKA, K. BEZDÌÈKOVÁ, K. HOLÝ, P. LIKA, J. MACEK, L. ROLLER, M. RÍHA et al. "Hymenoptera of Hády Hill, near the city of Brno (Czech Republic), collected during the Third Czech-Slovak Hymenoptera meeting." Acta Musei Moraviae, Scientiae biologicae (Brno) 93 (2008): 53-92.
- Ebsen J. R., J. J. Boomsma, and D. R. Nash. 2019. Phylogeography and cryptic speciation in the Myrmica scabrinodis Nylander, 1846 species complex (Hymenoptera: Formicidae), and their conservation implications. Insect Conservation and Diversity doi: 10.1111/icad.12366
- Elmes G. W. 1994. A population of the social parasite Myrmica hirsuta Elmes (Hymenoptera, Formicidae) recorded from Jutland, Denmark, with a first description of the worker caste. Insectes Soc. 41: 437-442.
- Elmes, G.W., T. Akino, J.A. Thomas, R.T. Clarke and J.J. Knapp. 2002. Interspecific differences in cuticular hydrocarbon profiles of Myrmica ants are sufficiently consistent to explain host specificity by Maculinea (large blue) butterflies. Oecologia 130:525-535
- Else G., B. Bolton, and G. Broad. 2016. Checklist of British and Irish Hymenoptera - aculeates (Apoidea, Chrysidoidea and Vespoidea). Biodiversity Data Journal 4: e8050. doi: 10.3897/BDJ.4.e8050
- Emery C. 1895. Beiträge zur Kenntniss der nordamerikanischen Ameisenfauna. (Schluss). Zoologische Jahrbücher. Abteilung für Systematik, Geographie und Biologie der Tiere 8: 257-360.
- Emery C. 1914. Wissenschaftliche Ergebnisse der Bearbeitung von O. Leonhard's Sammlungen. 5. Südeuropäische Ameisen (Hym.). Entomologische Mitteilungen. Berlin-Dahlem 3: 156-159.
- Emery C. 1916. Fauna entomologica italiana. I. Hymenoptera.-Formicidae. Bullettino della Società Entomologica Italiana 47: 79-275.
- Espadaler X., X. Roig, K. Gómez, and F. García. 2011. Formigues de les Planes de Son i mata de València (Hymenoptera, Formicidae) Treballs de la Institució Catalana d'Història Natural 16: 609-627.
- Espadaler, X., and C. Ascaso. "Adición a las hormigas del Montseny (Barcelona)." Orsis 5 (1990): 141-147.
- Espadaler, X. "Formicidos de las sierras de Cazorla, del Pozo y Segura (Jaén, España)." Ecología 11 (1997): 489-499.
- Espadaler, X. "Formigues del Montseny." in: Terradas, J., Miralles, J. (eds.) El patrimoni biològic del Montseny Catàlegs d (1987): 101-103.
- Fagan K. C., R. F. Pywell, J. M. Bullock, and R. H. Marrs. 2010. Are Ants Useful Indicators of Restoration Success in Temperate Grasslands? Restoration Ecology 18(3): 373379.
- Fiedler, K., F. Kuhlmann, B. C. Schlick-Steiner, F. M. Steiner and G. Gebauer. 2007. Stable N-isotope signatures of central European ants assessing positions in a trophic gradient. Insectes Sociaux 54(4):393-402.
- Finzi B. 1926. Le forme europee del genere Myrmica Latr. Primo contributo. Bollettino della Società Adriatica di Scienze Naturali in Trieste. 29: 71-119.
- Finzi, B. "Formiche dell'isola d'Elba e Monte Argentario." Bollettino della Società Entomologica Italiana 56 (1924): 12-15.
- Finzi, B. "Le forme europee del genere Myrmica Latr. Primo contributo." Bollettino della Società Adriatica di Scienze Naturali in Trieste 29 (1926): 71-119.
- Formidabel Database
- Fowles, A.P. 1996. A provisional checklist of the invertebrates recorded from Wales. 2. Aculeate wasps, bees and ants (Hymenoptera: Aculeata). Countryside Council for Wales
- Freitag A. 2013. Biodiversite 2010 en ville de Neuchatel: les fourmis (Hymenopteres, Formicidae). Bulletin de la societe Neuchateloise des sciences naturelles 133: 183-200.
- GRETIA. 2017. Bilan annuel de l'enquete sur la repartition des fourmis armoricaines. 23 pages.
- Gadeau de Kerville H. 1922. Materiaux pour la Faune des Hymenopteres de la Normandie. Bull. Soc. Amis Sc. Nat. Rouen 1916-1921, 1922: 217-225.
- Galkowski C. 2013. Nouvelles données sur la répartition de Strongylognathus huberi Forel, 1874 (Hymenoptera, Formicidae) en France. Bulletin de la Société Linnéenne de Bordeaux (n.s.) 41: 167-174.
- Galkowski C., and C. Foin. 2013. Nouvelles données sur la répartition de Strongylognathus huberi Forel, 1874 (Hymenoptera, Formicidae) en France. Bulletin de la Société Linnéenne de Bordeaux (n.s.) 41: 167-174.
- Galkowski C., and P Wegnez. 2010. Myrmica constricta Karavaiev 1934, nouvelle espece pour la France (Hymenoptera, Formicidae). Bull. Soc. Ent. Mulhouse 66(3): 41-45.
- Galle L. 1981. The Formicoid fauna of the Hortobagy. Pp. 307-311 in: Mahunka, S. (ed.) 1981. The fauna of the Hortobágy National Park. Budapest: Akadémiai Kiadó, 415 pp.
- Galle L. 1993. Data to the ant fauna of the Bukk (Hymenoptera: Formicidae). Natural history of the national parks of Hungary 7: 445-448.
- Galle L., and G. Szonyi. 1988. A check list of ants (Hymenoptera: Formicoidea) of a sandy grassland in Kiskunsag National Park (Hungary). Acta Biol. Szeged 34: 167-168.
- Gallé L. 1991. Structure and succession of ant assemblages in a north European sand dune area. Holarctic Ecology 14: 31-37.
- Gallé L., B. Markó, K. Kiss, E. Kovács, H. Dürgő, K. Kőváry, and S. Csősz. 2005. Ant fauna of Tisza river basin (Hymenoptera: Formicidae). In: Gallé, L. (szerk.): Vegetation and Fauna of Tisza River Basin I. Tiscia Monograph Series 7; Szeged, pp. 149-197.
- Gallé L., L. Körmöczi, E. Hornung, and J. Kerekes. 1998. Structure of ant assemblages in a Middle-European successional sand-dune area. Tiscia 31: 19-28.
- García F., X. Espadaler, P. Echave, and R. Vila. 2011. Hormigas (Hymenoptera, Formicidae) de los acantilados de l'Avenc de Tavertet (Osona) Boletín de la Sociedad Entomológica Aragonesa 47: 363-367.
- Gaspar C. 1968. Les fourmis de la Drome et des Basses-Alpes, en France (Hymenoptera, Formicidae). Naturaliste can. 95: 747-766.
- Gaspar C., and C. Thirion. 1978. Modification des populations d'Hymenopteres sociaux dans les milieux anthropogenes. Memorabilia Zoologica 29: 61-77.
- Gaspare Charles. 1965. Étude myrmécologique d'une région naturelle de Belgique: la Famenne Survey des Fourmis de la Région (Hymenoptera, Formicidae). Institut agronomique de l'Etat a' Gembloux. 32(4): 427-434.
- Ghahari H., and C. A. Collingwood. 2013. A study on the ants (Hymenoptera: Vespoidea: Formicidae) from Western Iran. Acta Phytopathologica et Entomologica Hungarica 48 (1): 155164.
- Giacalone I., and M. Moretti. 2001. Contributo alla conoscenza della mirmecofauna (Hymenoptera: Formicidae) dei castagneti al Sud delle Alpi (ticino, Svizzera). Bollettino della Societa ticinese di Scienze naturali 89(1-2): 51-60.
- Glaser F. 2009. Die Ameisen des Fürstentums Liechtenstein. (Hymenoptera, Formicidae). Amtlicher Lehrmittelverlag, Vaduz, 2009 (Naturkundliche Forschung im Fürstentum Liechtenstein; Bd. 26).
- Glaser F., A. Freitag, and H. Martz. 2012. Ants (Hymenoptera: Formicidae) in the Münstertal (Val Müstair) a hot spot of regional species richness between Italy and Switzerland. Gredleriana 12: 273 - 284.
- Glaser F., T. Kopf, and K. H. Steiberger. 2003. Ameisen (Hymenoptera: Formicidae) im Frastanzer Ried und den Illauen (Vorarlberg, Österreich) Artenspektrum, Gefährdung und Schutzempfehlungen. Vorarlberger Naturschau 13: 287-310.
- Gomez C., D. Casellas, J. Oliveras, and J. M. Bas. 2003. Structure of ground-foraging ant assemblages in relation to land-use change in the northwestern Mediterranean region. Biodiversity and Conservation 12: 21352146.
- Gomez K., D. Martinez, and X. Espadaler. 2018. Phylogeny of the ant genus Aphaenogaster (Hymenoptera: Formicidae) in the Iberian Peninsula, with the description of a new species. Sociobiology 65: 215-224.
- Gomez, C., P. Pons and J.M. Bas. 2003. Effects of the Argentine ant Linepithema humile on Seed Dispersal and Seedling Emergence of Rhamnus alaternus. Ecography 26(4):532-538
- Gouraud C. 2015. Bilan de l’année 2014 : Atlas des fourmis de Loire-Atlantique (Hymenoptera, Formicidae). Atlas des Formicidae de Loire-Atlantique, compte rendu de la première année d’étude (2014)
- Grandi G. 1935. Contributi alla conoscenza degli Imenotteri Aculeati. XV. Boll. R. Ist. Entomol. Univ. Studi Bologna 8: 27-121.
- Groc S., J. H. C. Delabie, R. Cereghino, J. Orivel, F. Jaladeau, J. Grangier, C. S. F. Mariano, and A. Dejean. 2007. Ant species diversity in the Grands Causses (Aveyron, France): In search of sampling methods adapted to temperate climates. C. R. Biologies 330: 913922.
- Grzes I. M. 2009. Ant species richness and evenness increase along a metal pollution gradient in the Boles?aw zinc smelter area. Pedobiologia 53: 65-73.
- Haguet G., M. Chevrier, and E. Brunel. 2002. Les invertebres de la dune de Bon Abri, premier inventaire. Groupe d'Etude des Invertebres Armoricains, 37 pages.
- Hauschteck-Jungen E., and H. Jungen. 1983. Ant chromosomes. II. Karyotypes of western palearctic species. Insectes Soc. 30: 149-164.
- Holec M., and J. Frouz. 2005. Ant (Hymenoptera: Formicidae) communities in reclaimed and unreclaimed brown coal mining spoil dumps in the Czech Republic. Pedobiologia 49: 345-357.
- Holecova M., M. Klesniakova, K. Holla, and A. Sestakova. 2017. Winter activity in scots pine canopies in Borska Nizina Lowland (SW Slovakia)
- Holgersen H. 1940. Myrmekologiske notiser I. Nor. Entomol. Tidsskr. 5: 183-187.
- Holgersen H. 1943. Ant studies in Rogaland (south-western Norway). Avhandlingar utgitt av det Norske Videnskaps-Akademi i Oslo. I. Matematisk-Naturvidenskapelig Klasse 1943(7): 1-75.
- Holgersen H. 1944. The ants of Norway (Hymenoptera, Formicidae). Nytt Magasin for Naturvidenskapene 84: 165-203.
- Holgersen H. 1944. The ants of Norway (Hymenoptera, Formicidae). Nytt Magasin for Naturvidenskapene 84: 165-450.
- Hughes J. 1994. Notable records of ants (Hymenoptera: Formicidae) in south-east Sutherland. Entomologist's Record and Journal of Variation 106: 75-76.
- IZIKO South Africa Museum Collection
- Jansen G., R. Savolainen, K. Vespalainen. 2010. Phylogeny, divergence-time estimation, biogeography and social parasite–host relationships of the Holarctic ant genusMyrmica(Hymenoptera: Formicidae). Molecular Phylogenetics and Evolution 56: 294-304.
- Jeffery H. G. 1931. The Formicidae (or ants) of the Isle of Wight. Proceedings of the Isle of Wight Natural History and Archaeological Society 2: 125-128.
- Kanizsai O., R. Gallé, and L. Gallé. 2009. Perception of spatial patchiness by ant assemblages (Hymenoptera: Formicidae). Tiscia 37: 3-7.
- Karaman M. G. 2011. A catalogue of the ants (Hymenoptera, Formicidae) of Montenegro. Podgorica: Catalogues 3, Volume 2, Montenegrin Academy of Sciences and Arts, 140 pp.
- Karaman M. G., and G. S. Karaman. 2007. Contribution to the Knowledge of the ants (Hymenoptera, Formicidae) from Special nature reserve Zasavica , Serbia. Zbornik “Naucno-strucni skup Zasavica 2007, Sremska Mitrovica, 67-75.
- Kiran K., and C. Karaman. 2012. First annotated checklist of the ant fauna of Turkey (Hymenoptera: Formicidae). Zootaxa 3548: 1-38.
- Kiran K., and N. Aktac. 2006. The vertical distribution of the ant fauna (Hymenoptera: Formicidae) of the Samanh Mountains, Turkey. Linzer Biol. Beitr. 38(2): 1105-1122.
- Kofler A. 1995. Nachtrag zur Ameisenfauna Osttirols (Tirol, Österreich) (Hymenoptera: Formicidae). Myrmecologische Nachrichten 1: 14-25.
- Krausse, A. H. "Ueber sardische Ameisen." Archiv für Naturgeschichte, Berlin A 78(7) (1912): 162-166.
- Kvamme T. 1982. Atlas of the Formicidae of Norway (Hymenoptera: Aculeata). Insecta Norvegiae 2: 1-56.
- Kvamme T. 1999. Notes on Norwegian ants (Hymenoptera, Formicidae). Norwegian Journal of Entomology 46: 19-23.
- Lapeva-Gjonova, L., V. Antonova, A. G. Radchenko, and M. Atanasova. "Catalogue of the ants (Hymenoptera, Formicidae) of Bulgaria." ZooKeys 62 (2010): 1-124.
- Le Moli F., and A. Zaccone. 1995. Ricerche sulla mirmecofauna del Cansiglio (Prealpi Carniche). Soc. Ven. Sc. Nat. 20: 33-52.
- Le Moli F., and M. R. Rose. 1991. Atti della Societa Italiana di Societa Italiana di Scienze Naturali e del Museo Civico di Storia Naturale di Milan. Atti della Società Italiana di Scienze Naturali e del Museo Civico di Storia Naturale di Milano 132:25-40.
- Lebas C., C. Galkowski, P. Wegnez, X. Espadaler, and R. Blatrix. 2015. The exceptional diversity of ants on mount Coronat (Pyrénées-Orientales), and Temnothorax gredosi(Hymenoptera, Formicidae) new to France. R.A.R.E., T. XXIV (1): 24 33
- Lebas C., and C. Galkowski. 2016. Myrmica hirsuta Elmes, 1978, a new species from France (Hymenoptera, Formicidae). Bull. Soc. Linn. Bordeaux 151, 44(2/3): 239-244.
- Legakis Collection Database
- Lehouck V., D. Bonte, W. Dekoninck, and J. P. Maelfait. 2004. The distribution of ant nests (Hymenoptera, Formicidae) in coastal grey dunes of Flanders (Belgium) and their relationship to myrmecochorous plants. Belg. J. Zool. 134 (2/1) : 89-96.
- Lenoir A. 1971. Les fourmis de Touraine, leur intérêt biogéographique. Cahiers des Naturalistes 27: 21-30.
- Lillig M., and E. Dewes. 2015. The former Siegfried Line as habitats for ants (Hymenoptera: Formicidae). Abh. Delattinia 37: 117 - 136
- Livory A. 2003. Les fourmis de la Manche. L'Argiope 39: 25-49.
- Livory A. Un Nouvel Inventaire: les Fourmis. Argiope 29: 27-34.
- Lorinczi G. 2011. Density and spatial pattern of nests in sub-Mediterranean ground-dwelling ant communities (Hymenoptera: Formicidae). Community Ecology 12(1): 51-57.
- Majzlan O., and P. Devan. 2009. Selected insect groups (Hymenoptera, Neuroptera, Mecoptera, Raphidioptera) of the Rokoš Massif (Strážovské vrchy Mts.). Rosalia (Nitra), 20, p. 63–70.
- Mann W. M. 1911. On some Northwestern ants and their guests. Psyche (Cambridge) 18: 102-109.
- Marko B. 2008. Ants (Hymenoptera: Formicidae) of the Sand Dunes of Foieni Protected Area and Its Surroundings (Satu Mare County, Romania), and a New Species for the Romanian Fauna. Acta Scientiarum Transylvanica 16(3): 87-99.
- Markó B., B. Sipos, S. Csősz, K. Kiss, I. Boros, and L. Gallé. 2006. A comprehensive list of the ants of Romania (Hymenoptera: Formicidae). Myrmecologische Nachrichten 9: 65-76.
- Markó B., and S. Csősz. 2002. Die europäischen Ameisenarten (Hymenoptera: Formicidae) des Hermannstädter (Sibiu, Rumänien) Naturkundemuseums I.: Unterfamilien Ponerinae, Myrmicinae und Dolichoderinae. Annales Historico-Naturales Musei Nationalis Hungarici 94: 109-121.
- Mei, M. "Myrmica samnitica n. sp.: una nuova formica parassita dell'Appennino Abruzzese (Hymenoptera, Formicidae)." Fragmenta Entomologica 19 (1987): 457-469.
- Meyer M., C. Braunert, R. Gerend, and I. Schrankel. 2003. Résultats de l'excursion annuelle du groupe de travail entomologique de la Société des naturalistes luxembourgeois. Bull. Soc. Nat. luxemb. 103: 103-120.
- Monteserín Real, S. "Invertebrados de la Reserva Natural Integral de Muniellos, Asturias: Formicidae." Consejería de Medio Ambiente, Ordenación del Territorio e Infraestructuras del Principado de Asturias KRK edicio (2003): 269.
- Moretti M. P. Duelli, and M. K. Obrist. 2006. Biodiversity and resilience of arthropod communities after fire disturbance in temperate forests. Oecologia 149: 312327.
- Mouquet C. 2006. Premier inventaire des invertébrés terrestres des plages du département du Calvados. Rapport Gretia pour le Syndicat Mixte Calvados Littoral Espaces Naturels. 36 p.
- Müller, G. "Le formiche della Venezia Guilia e della Dalmazia." Bollettino della Società Adriatica di Scienze Naturali in Trieste 28 (1923): 11-180.
- Nemet E., Z. Czekes, I. Tausan, and B. Marko. 2012. Contribution to the knowledge of the myrmecofauna of the Cefa Nature Park (North-Western Romania). Acta Scientiarum Transylvanica 20(1): 61-72.
- Neumeyer R., and B. Seifert. 2005. Commented check list of free living ant (Hymenoptera: Formicidae) species of Switzerland. Bulletin de la Societe Entomologique Suisse 78: 1-17.
- O'Rourke F. J. 1948. The distribution and general ecology of the Irish Formicidae. Proceedings of the Royal Irish Academy. Section B: Biological, Geological, and Chemical Science 52: 383-410.
- Odegaard F. 2013. New and little known ants (Hymenoptera, Formicidae) in Norway. Norwegian Journal of Entomology 60, 172175.
- Ottonetti L., L. Tucci, and G. Santini. 2006. Recolonization Patterns of Ants in a Rehabilitated Lignite Mine in Central Italy: Potential for the Use of Mediterranean Ants as Indicators of Restoration Processes. Restoration Ecology 14(1): 6066.
- Palanca, A., A. Urbieta, C. Castán, and X. Espadaler. "Ants in the supraforestal pastures of the Tendeñera massif." Écologie des milieux montagnards et de haute altitude. Documents d'Ecologie Pyrénéenne III-IV (1984): 223-224.
- Paraschivescu D. 1978. Elemente balcanice in mirmecofauna R. S. Romania. Nymphaea 6: 463- 474.
- Petal J. 1974. Analysis of a sheep pasture ecosystem in the Pieniny mountains (the Carpathians) XV. The effect of pasture management on ant population. Ekologia Polska 22(3/4): 679-692.
- Petrov I. Z. 1986. Contribution to myrmecofauna in some oak-tree communities on the mountain Jastrebac. Prirodnjackog Muzeja i Beogradu Seriya B Bioloske Nauke Supplement: No. 41: 109-114.
- Petrov I. Z. 2002. Contribution to the myrmecofauna (Formicidae, Hymenoptera) of the Banat Province (Serbia). Archives of Biological Sciences, Belgrade, 54(12): 57-64.
- Petrov I. Z., B. Petrov, D. Milicic, T. Karan-Znidarsic. 2007. Contribution to the Myrmecofauna (Hymenoptera: Formicidae) of East and South Serbia. Acta Zoologica Bulgarica 59(3): 295-299.
- Petrov I. Z., and C. A. Collingwood. 1992. Survey of the myrmecofauna (Formicidae, Hymenoptera) of Yugoslavia. Archives of Biological Sciences (Belgrade) 44: 79-91.
- Pisarski B. 1982. Ants (Hymenoptera: Formicoidea) of Warsaw and Mazovia. Memorabilia Zool. 36: 73-90.
- Poldi B. 1980. Imenotteri formicidi della Brughiera di Rovasenda (Piemonte). In: Poldi, B. 1980. Quaderni sulla "Struttura delle zoocenosi terrestri". 1. La brughiera pedemontana. III. Serie AQ/1/110. Collana del programma finalizzato "Promozione della qualità dell'ambiente". Roma: Consiglio Nazionale delle Ricerche, pp. 7-19.
- Poldi B., M. Mei, and F. Rigato. 1995. Hymenoptera, Formicidae. Vol 102. Checklist delle specie della fauna Italiana: 1-10.
- Pusvaskyte O. 1979. Myrmecofauna of the Lituanian SSR. Acta Entomologica Lituanica 4: 99-105.
- Retana J., and X. Cerdá X. 2000. Patterns of diversity and composition of Mediterranean ground ant communites tracking spatial and temporal variability in the thermal environment. Oecologia 123: 436-444.
- Rigato F., and R. Sciaky. 1989. Contributo alla conoscenza della mirmecofauna della Val Gesso (alpi Marittime) (Hymenoptera, Formicidae). Boll. Mus. Reg. Sci. Nat. Torino 7(2): 427-442.
- Rigato F., and R. Sciaky. 1991. The myrmecofauna of the Gesso Valley (Maritime Alps) (Hymenoptera Formicidae). Ethology Ecology and Evolution Special Issue 1: 87-89.
- Saaristo M. I. 1990. New provincial records for twelve ant species (Hymenoptera, Formicidae) from Finland. Entomologica Fennica 1: 191-192.
- Sadil J. V. 1952. A revision of the Czechoslovak forms of the genus Myrmica Latr. (Hym.). Sb. Entomol. Oddel. Nár. Mus. Praze 27: 233-278.
- Salata S. 2014. Ants (Hymenoptera: Formicidae) of the National Park of the Stołowe Mts. Przyroda Sudetow 17: 161-172.
- Salata S., and L. Borowiec. 2018. Taxonomic and faunistic notes on Greek ants (Hymenoptera: Formicidae). Annals of the Upper Silesian Museum in Bytom Entomology 27: 1-51.
- Sanders, D. and C. Platner. 2007. Intraguild Interactions between Spiders and Ants and Top-Down Control in a Grassland Food Web. Oecologia 150(4):611-624
- Saure C. 2005. Rote Liste und Gesamtartenliste der Bienen und Wespen (Hymenoptera part.) von Berlin mit Angaben zu den Ameisen. In: DER LANDESBEAUFTRAGTE FÜR NATURSCHUTZ UND LANDSCHAFTSPFLEGE / SENATSVERWALTUNG FÜR STADTENTWICKLUNG (Hrsg.): Rote Listen der gefährdeten Pflanzen und Tiere von Berlin. CD-ROM.
- Schlick-Steiner B. C., and F. M. Steiner. 1999. Faunistisch-ökologische Untersuchungen an den freilebenden Ameisen (Hymenoptera: Formicidae) Wiens. Myrmecologische Nachrichten 3: 9-53.
- Schlick-Steiner, B.C., F.M. Steiner, K. Moder, A. Bruckner, K. Fiedler and E. Christian. 2006. Assessing ant assemblages: pitfall trapping versus nest counting (Hymenoptera, Formicidae). Insectes Sociaux 53:274-281
- Seifert B. 1988. A taxonomic revision of the Myrmica species of Europe, Asia Minor, and Caucasia (Hymenoptera, Formicidae). Abhandlungen und Berichte des Naturkundemuseums Görlitz 62(3): 1-75.
- Seifert B. 1994. Die freilebenden Ameisenarten Deutschlands (Hymenoptera: Formicidae) und Angaben zu deren Taxonomie und Verbreitung. Abhandlungen und Berichte des Naturkundemuseums Görlitz 67(3): 1-44.
- Seifert, B. "A taxonomic revision of the Myrmica species of Europe, Asia Minor, and Caucasia." Abhandlungen und Berichte des Naturkundemuseums Goerlitz 62(3) (1988): 1-75.
- Sellier Y., C. Galkowski, C. Lebas, and P. Wegnez. 2016. Découverte de Temnothorax pardoi (Tinaut, 1987) dans la réserve naturelle nationale du Pinail (Hymenoptera, Formicidae). Revue de l’Association Roussillonnaise d’Entomologie 25(2): 106-113.
- Sielezniew M., D. Patricelli, I. Dziekanska, F. Barbero, S. Bonelli, L. P. Casacci, M. Witek, and E. Balletto. 2010. The frst record of Myrmica lonae (Hymenoptera: Formicidae) as a host of the socially parasitic large blue butterfly Phengaris (Maculinea) arion (Lepidoptera: Lycaenidae). Sociobiology 56(2): 465-.
- Smith F. 1941. A list of the ants of Washington State. The Pan-Pacific Entomologist 17(1): 23-28.
- Somogyi A. A., G. Lorinczi, J. Kovacs, and I. E. Maak. 2017. Structure of ant assemblages in planted poplar (Populus alba) forests and the effect of the common milkweek (Asclepias syriaca). Acta Zoologica Academiae Scientiarum Hungaricae 63(4): 443–457.
- Sonnenburg H. 2005. Die Ameisenfauna (Hymenoptera: Formicidae) Niedersachsens und Bremens. Braunschweiger Naturkundliche Schriften 7: 377-441.
- Soudek, S. "Dalmatsti mravenci." Casopis Csl. Spolecnosti Entomologické 20 (1-2) (1925): 12-17.
- Stankiewicz A. M., M. Sielezniew, and G. Svitra. 2005. Myrmica schencki (Hymenoptera: Formicidae) rears Maculinea rebeli (Lepidoptera: Lycaenidae) in Lithuania: new evidence for geographical variation of host-ant specificity of an endangered butterfly. Myrmecologische Nachrichten 7: 51-54.
- Steiner F. M., S. Schödl, and B. C. Schlick-Steiner. 2002. Liste der Ameisen Österreichs (Hymenoptera: Formicidae), Stand Oktober 2002. Beiträge zur Entomofaunistik 3: 17-25.
- Stukalyuk S. V. 2015. Structure of the ant assemblages (Hymenoptera, Formicidae) in the broad-leaved forests of Kiev. Entomological Review 95(3): 370–387.
- Stukalyuk S. V. 2017. Stratification of the ant species (Hymenoptera, Formicidae) in the urban broadleaf woodlands of the city of Kiev. Entomological Review 97(3): 320-343.
- Sturtevant A. H. 1931. Ants collected on Cape Cod, Massachusetts. Psyche (Cambridge) 38: 73-79
- Suñer i Escriche, David. "Contribució al coneixement mirmecologic de Gavarres, Montgrí, Guilleríes i la Serralada Transversal." Tesis Doctoral Universida (1991): 577 pp.
- Szujecki A., J. Szyszko, S. Mazur, and S. Perlinski. 1978. A succession of the ants (Formicidae) on afforested arable land and forest soils. Memorabilia Zoologica 29: 183-189.
- Tartally, A., D.R. Nash, S. Lengyel and Z. Varga. 2008. Patterns of host ant use by sympatric populations of Maculinea alcon and M. rebeli in the Carpathian Basin. Insectes Sociaux 55:370-381
- Tausan I., M. M. Jerpel, I. R. Puscasu, C. Sadeanu, R. E. Brutatu, L. A. Radutiu, and V. Giurescu. 2012. Ant fauna (Hymenoptera: Formicidae) of Sibiu County (Transylvania, Romania). Brukenthal. Acta Musei 7(3): 499-520.
- Ticha, K. 2005. Inventarizacní pruzkum mravencu (Hymenoptera: Formicidae) NPP Svarec. 2005. Acta Rerum Naturalium 1:127-130.
- Tinaut A. 2016. Ants of the Tejeda, Almijara and Alhama Mountains Natural Park (Andalusia, Spain) (Hymenoptera, Formicidae). Boln. Asoc. esp. Ent., 40 (1-2): 125-159.
- Urbieta Balado A., A. Palanca Soler, C. Castan Lanaspa, and X. Espadaler. 1984. Ants in the supraforestal pastures of the Tendenera massif. Documents dEcologie Pyrénéenne III-IV: 223-224.
- Vagalinski B., and A. Lapeva-Gjonova. 2012. The ants (Hymenoptera, Formicidae) of Plana Mountain (Bulgaria). Historia naturalis bulgarica 20: 87-101.
- Vassilev I. 1987. The ants (Formicidae, Hymenoptera) from the valley of the river Racene. Annuaire de l'Universite de Sofia Kliment Ohridski Faceulte de Biologie 78: 74-79.
- Vepsalainen K., H. Ikonene, and M. J. Koivula. 2008. The structure of ant assembalges in an urban area of Helsinki, southern Finland. Ann. Zool. Fennici 45: 109-127.
- Verdcourt, B. 2004. Additions to the Wild Fauna and Flora of the Royal Botanic Gardens, Kew XXXV. Miscellaneous Records. Kew Bulletin 59(4):639-649
- Versluijs R., and P. Boer. 2014. Socially parasitic Myrmicaants of The Netherlands, including a new species: Myrmica bibikoffi(Hymenoptera: Formicidae). Entomologische Berichten 74 (4): 147-151.
- Vesni? A. 2011: Revidirani sistematski prijegled mrava Bosne i Hercegovine. Unutar : S. Lelo (urednik), Fauna Bosne i Hercegovine Biosistematski pregledi. 7. izmijenjeno i popravljeno interno izdanje Udruenja za inventarizaciju i zatitu ivotinja, Ilija, Kanton Sarajevo, pp: 205-207.
- Wegnez P. 2014. Premières captures de Lasius distinguendus Emery, 1916 et de Temnothorax albipennis (Curtis, 1854) au Grand-Duché de Luxembourg (Hymenoptera : Formicidae). Bulletin de la Société royale belge d’Entomologie 150 (2014) : 168-171.
- Wegnez P. 2017. Découverte de Myrmica lobicornis Nylander, 1846 et Lasius jensi Seifert, 1982, deux nouvelles espèces pour le Grand-Duché de Luxembourg (Hymenoptera: Formicidae). Bulletin de la Société royale belge d’Entomologie 153: 46-49.
- Wegnez P. 2018. Premières decouvertes de Myrmica bibikoffi Kutter, 1963 et de Ponera testacea Emery, 1895, au Luxembourg (Hymenoptera: Formicidae)Bulletin de la Société royale belge d’Entomologie 154: 263–272.
- Wegnez P., D. Ignace, E. Lommelen, M. Hardy, J. Bogaert, and C. Nilsson. 2015. Redécouverte de Teleutomyrmex schneideriKutter, 1950 dans les Alpes françaises (Hymenoptera: Formicidae). Bulletin de la Société royale belge d’Entomologie 151: 52-57.
- Wegnez P., and A. Ronk. 2017. Découverte de Camponotus herculeanus (Linnaeus, 1758) et signalement de quelques autres espèces rares de fourmis au Luxembourg (Hymenoptera : Formicidae). Bulletin de la Société des naturalistes luxembourgeois 119 : 153–159.
- Wegnez P., and M. Fichaux. 2015. Liste actualisee des especes de fourmis repertoriees au Grand-Duche de Luxembourg (Hymenoptera: Formicidae). Bulletin de la Société royale belge d’Entomologie 151: 150-165
- Wheeler W. M. 1917. A list of Indiana ants. Proceedings of the Indiana Academy of Science 26: 460-466.
- Wheeler W. M. 1928. Ants of Nantucket Island, Mass. Psyche (Cambridge) 35: 10-11.
- Wiezik M. 2006. Ants (Hymenoptera: Formiciadae) of the Sitno National Nature Reserve (tiavnické vrchy Protected Landscape Area). Naturae Tutela 10: 73-77.
- Wiezik M., A. Wiezikova, and M. Svitok. 2010. Effects of secondary succession in abandoned grassland on the activity of ground-foraging ant assemblages (Hymenoptera: Formicidae). Acta Soc. Zool. Bohem. 74: 153-160
- Wieziková A. 2008. Epigaeic activity of ants (Hymenoptera: Formicidae) at hemerobic gradient: arable land green forage permanent grassland. Acta Facultatis Ecologiae 18: 69-76.
- Wlodarczyk T. 2010. Ant species composition in relation to forest associations in Szczecin Landscape Park. Polish Journal of Entomology 79: 91-99.
- Wlodarczyk T., M. Zmihorski, and A. Olczyk. 2009. Ants inhabiting stumps on clearcuts in managed forest in western Poland. Entomologia Fennica 20: 121-128.
- Zimmermann, S. "Beitrag zur Kenntnis der Ameisenfauna Süddalmatiens." Verhandlungen der Zoologisch-Botanisch Gesellschaft in Wien 84 (1935): 1-65.
- de Biseau, J.-C., Y. Quinet, L. Deffernez and J.M. Pasteels. 1997. Explosive food recruitment as a competitive strategy in the ant Myrmica sabuleti (Hymenoptera, Formicidae). Insectes Sociaux 44:59-73
- de Haro, Andrés, and C. A. Collingwood. "Prospección mirmecológica en la Cordillera Ibérica." Orsis 6 (1991): 129-126.
- Pages using DynamicPageList3 parser function
- Polygynous
- North temperate
- FlightMonth
- Ant Associate
- Host of Myrmica bibikoffi
- Host of Myrmica hirsuta
- Host of Myrmica karavajevi
- Butterfly Associate
- Host of Maculinea arion
- Host of Myrmica laurae
- Host of Phengaris arion
- Nematode Associate
- Host of Mermithidae (unspec.)
- Host of Hexamermis/Agamermis
- Fungus Associate
- Host of Hormiscium myrmecophilum
- Host of Rickia wasmanniii
- Host of Rickia wasmannii
- Karyotype
- Species
- Extant species
- Formicidae
- Myrmicinae
- Myrmicini
- Myrmica
- Myrmica sabuleti
- Myrmicinae species
- Myrmicini species
- Myrmica species
- Need Body Text