Pachycondyla striata
| Pachycondyla striata | |
|---|---|

| |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Arthropoda |
| Class: | Insecta |
| Order: | Hymenoptera |
| Family: | Formicidae |
| Subfamily: | Ponerinae |
| Tribe: | Ponerini |
| Genus: | Pachycondyla |
| Species: | P. striata |
| Binomial name | |
| Pachycondyla striata Smith, F., 1858
| |
| Synonyms | |
| |


{kind=link}
Pachycondyla striata nests in rotten wood, under logs, or in the soil and forages on the soil surface (Mackay and Mackay).
Identification
From Mackay and Mackay (2010): There are several species of New World Pachycondyla in which the workers and females have conspicuous striae on the dorsum of the head, including Paltothyreus tarsatus, Pachycondyla fuscoatra, Neoponera lineaticeps, Neoponera magnifica and P. striata. Pachycondyla striata can be easily separated from P. tarsata, as it lacks the inner claw on the tarsal claws and the swellings on the upper anterior region of the postpetiole, both of which are present in P. tarsata. Pachycondyla striata differs from N. lineaticeps in the shape of the petiole, which is nearly rectangular, not with the posterior face broadly rounded into the dorsal face. Additionally P. striata is much larger (total length of N. lineaticeps 10 mm). Pachycondyla striata can also be separated as it occurs mostly in southern South America, not Central America and northern South America as does N. lineaticeps. The striae on the head and dorsum of the pronotum of P. striata are not as coarse and well defined as they are in N. magnifica. Pachycondyla striata has approximately 50 striae across the widest point of the pronotum, whereas N. magnifica has about 35. Pachycondyla striata could be easily confused with P. fuscoatra. It differs in having a well-developed pronotal carina (lacking in P. fuscoatra) and the stria on the head are finer and not as well defined. Pachycondyla striata can be confused with Pachycondyla impressa, but differs in that P. impressa lacks the striae on the head and lacks a well-developed carina on the pronotal shoulder. Otherwise they are very similar.
The males of three common species are very similar: P. striata, P. impressa and Neoponera villosa. Pachycondyla striata can be separated from the other two by the presence of dense erect hairs on most of the surfaces (sparse or only moderately abundant in the other two species). The subpetiolar process of P. striata is angulate anteriorly, not broadly rounded as in P. impressa, or with a large ventral process in N. villosa.
Keys including this Species
Distribution
BRASIL, BOLIVIA, PARAGUAY, ARGENTINA, URUGUAY (Mackay and Mackay 2010)
Latitudinal Distribution Pattern
Latitudinal Range: 27.158° to -64.36°.
| North Temperate |
North Subtropical |
Tropical | South Subtropical |
South Temperate |
- Source: AntMaps
Distribution based on Regional Taxon Lists
Neotropical Region: Argentina, Brazil (type locality), Paraguay, Uruguay.
Distribution based on AntMaps
Distribution based on AntWeb specimens
Check data from AntWeb
Countries Occupied
| Number of countries occupied by this species based on AntWiki Regional Taxon Lists. In general, fewer countries occupied indicates a narrower range, while more countries indicates a more widespread species. |

|
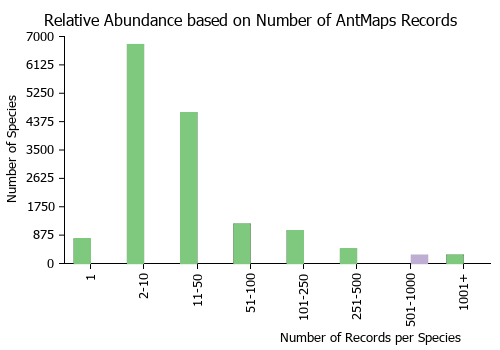
Estimated Abundance
| Relative abundance based on number of AntMaps records per species (this species within the purple bar). Fewer records (to the left) indicates a less abundant/encountered species while more records (to the right) indicates more abundant/encountered species. |
|
Habitat
Pachycondyla striata can be found in wet myrtaceous forest, rain forest, riparian rain forest, open grassy areas and cerrado [scrub vegetation], at sea level to 1300 meters elevation. (Mackay and Mackay 2010)
Biology
Nests of P. striata are simple and poorly elaborated, with the chambers and tunnels horizontally distributed close to the surface of the soil (Silva-Melo et al. 2010). Established colonies (N=7) contained 150 - 384 workers.
Rosumek (2017) examined trophic and temporal niches of ants in a secondary Atlantic forest (Florianópolis, south Brazil; 27°31′38″S 48°30′15″W / 27.52722°S 48.50417°W), between December 2015 and January 2016. For this species: Odontomachus chelifer and Pachycondyla striata are two widespread species that differ radically in morphology but are similar in many aspects. It is appropriate to discuss both together. They are one of the most conspicuous elements of the southern Atlantic forest ground fauna, due to their abundance, solitary foraging mode and large size. Several accounts show these species occupy a multitude of functional roles and have a broad trophic niche. Observation of nest entrances showed that 80-90% of the items carried by P. striata were arthropod parts, mostly termites and other ants, with the remainder consisting of plant material (Giannotti & Machado, 1991; Medeiros & Oliveira, 2009). Through direct observation, Medeiros and Oliveira (2009) also showed that scavenging accounts for more than 80% of its foraging behavior. On the other hand, Fowler (1980) reported O. chelifer prefers certain termite species in the laboratory, and qualitatively stated that in the field prey consisted almost entirely of termites. In the Atlantic forest, all items carried to nests were arthropods, mainly termites, but other animal groups accounted for 60% of them (Raimundo et al., 2009). Scavenging was also cited in this study, although not quantified. Both O. chelifer and P. striata were recorded on experimental carcasses predating the larvae and pupae of necrophagous insects (Paula et al., 2016).
Other important items used by the two species are seeds with elaiosomes and other fallen diaspores rich in proteins and lipids, frequently collected from the ground in the Atlantic forest (Pizo & Oliveira, 2000; Passos & Oliveira, 2002, 2004). Field records on use of liquid sugars are scant and qualitative. Odontomachus chelifer was not observed using extra-floral nectaries by Raimundo et al. (2009), while there is one record for P. striata (in Cerrado, the Brazilian savannah – Byk & Del-Claro, 2010).
The results presented here are mostly consistent with this broader picture. Both species more frequently foraged upon dead insects, sucrose and large prey. In fact, they were the only two species consistently recorded on crickets. The low frequency in termite baits is unexpected and probably represents a methodological artifact. These large solitary foragers were observed quickly collecting termites (even glued ones) and leaving the baits in a few minutes, contrary to ants of smaller species that recruited to them. In cricket baits, however, the two species spent more time trying to carry out the tied cricket or dismembering it. Smaller ants frequently took advantage of this to grab the remains or lick spilled hemolymph. This largely contributed to the richness found in this bait and could happen in nature, whenever predators kill prey too large to carry them out at once.
Besides predation, scavenging was a common behavior, and P. striata in particular would prioritize it whenever possible, in accordance with what was observed by Medeiros and Oliveira (2009). This could be result of their morphology, because the triangular mandibles may be more suited to chop large carcasses than the trap-jaws of O. chelifer. In addition, it could be an effect of competition. The two species were never found at the same bait, and co-occurred in just 10 of the 62 points where they were recorded. The two were previously reported to avoid each other, but, when agonistic interactions occur, P. striata usually is the winner, and can steal the food or kill (and eat) O. chelifer (Medeiros & Oliveira, 2009; Raimundo et al., 2009). Thus, P. striata could displace O. chelifer and maintain control of a valuable resource such as dead arthropods through tandem recruitment (Medeiros & Oliveira, 2009; Silva-Melo & Giannotti, 2012), while cooperative foraging behavior was not observed in O. chelifer (Raimundo et al., 2009). Effectively, the average numbers of workers per bait was smaller for the latter species (O. chelifer = 1.4 ± 0.9; P. striata = 2.5 ± 2; Mann-Whitney, z = -2.27, p = 0.02).
Both species used sucrose frequently and rarely foraged on melezitose (anolog for honeydew). They were never observed foraging on trees or low vegetation in this study, which fits previous accounts (Fowler, 1980; Medeiros & Oliveira 2009), so it is unlikely that they commonly use nectar as food source. This behavior also should limit honeydew use by them.
Past results have shown that these species show a preference for day (P. striata) or night foraging (O. chelifer). No preference in their time of foraging was found in this study.
Venom
Santos et al. (2017) - Pachycondyla striata has a distinguished behavior, producing a foamy substance from terminal abdominal region, as a defense mechanism. The venom, released as a foam-like substance (Mackay & Mackay, 2010), has been also reported in Pseudoneoponera tridentata and Pseudoneoponera insularis (both of the former were originally described as Pachycondyle species) (Maschwitz et al., 1981). However the venom compounds in this ant remain unknown. Thus, the objective of this study was evaluate the protein composition of the venom in the predatory ant P. striata collected in the state of Minas Gerais, Brazil.
The venom of P. striata has a complex mixture of proteins from which 43 were identified. Within the identified proteins are classical venom proteins (phospholipase A, hyaluronidase, and aminopeptidase N), allergenic proteins (different venom allergens), and bioactive peptides (U10-ctenitoxin Pn1a). Venom allergens are among the most expressed proteins, suggesting that P. striata venom has high allergenic potential. This study discusses the possible functions of the proteins identified in the venom of P. striata.
Association with Other Organisms
 Explore: Show all Associate data or Search these data. See also a list of all data tables or learn how data is managed.
Explore: Show all Associate data or Search these data. See also a list of all data tables or learn how data is managed.
Diptera
- This species is a host for the phorid fly Apocephalus piliventris (a parasite) (Brown et al., 2015) (injured).
- This species is a host for the phorid fly Apocephalus piliventris (a parasite) (phorid.net) (attacked).
Nematode
- This species is a host for the nematode Mermithidae (Mermithidae (unspecified "Mermis") (a parasite) in Paraguay (Strelnikov, 1928; Laciny, 2021).
Fungi
- This species is a host for the fungus Ophiocordyceps australis (a pathogen) (Shrestha et al., 2017).
Castes
Queen
Images from AntWeb
    
| |
| Queen (alate/dealate). Specimen code casent0178186. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by ALWC, Alex L. Wild Collection. |
Nomenclature
The following information is derived from Barry Bolton's Online Catalogue of the Ants of the World.
- striata. Pachycondyla striata Smith, F. 1858b: 106 (w.) BRAZIL (Rio de Janeiro).
- Type-material: 6 syntype workers.
- Type-locality: Brazil: Rio de Janeiro, “Rio 57/63” (H. Clark).
- Type-depositories BMNH, OXUM.
- [Note: 5 worker syntypes BMNH and 1 worker syntype OXUM. BMNH Accessions Register has: “1857 no. 63 (June 22). Province of Rio. Presented by Rev. Hamlet Clark.” Three more workers in BMNH are mounted in the same way as the syntypes but have passed through the Farren White collection and had their data labels altered to read “Rio. H. Clark.” These are probably also syntypes.]
- Roger, 1861a: 6 (q.); Santschi, 1921g: 86 (m.); Wheeler, G.C. & Wheeler, J. 1952c: 616 (l.).
- Combination in Ponera (Pachycondyla): Roger, 1861a: 6;
- combination in Pachycondyla (Pachycondyla): Emery, 1901a: 45;
- combination in Pachycondyla: Mayr, 1863: 439; Schmidt, C.A. & Shattuck, 2014: 155.
- Status as species: Roger, 1861a: 6; Mayr, 1862: 720; Roger, 1863b: 18; Mayr, 1863: 439; Mayr, 1865: 65; Emery, 1878a: ix (in list); Emery, 1888c: 353; Emery, 1890a: 71 (in key); Dalla Torre, 1893: 35; von Jhering, 1894: 380; Forel, 1895b: 115; Emery, 1896h: 625; Emery, 1906c: 114; Forel, 1906d: 249Forel, 1907a: 6; Forel, 1907e: 1; Forel, 1908c: 345; Emery, 1911d: 75; Forel, 1911e: 254; Bruch, 1914: 214; Santschi, 1916e: 366; Gallardo, 1918b: 62 (redescription); Luederwaldt, 1918: 35; Santschi, 1919f: 39; Santschi, 1921g: 86; Borgmeier, 1923: 70; Wheeler, W.M. 1925a: 5; Santschi, 1929d: 276; Eidmann, 1936a: 34; Kusnezov, 1953b: 336; Kempf, 1961c: 201; Kempf, 1964e: 52 (in key); Kempf, 1972a: 175; Kempf & Lenko, 1976: 52; Zolessi, et al. 1988: 2; Bolton, 1995b: 310; Wild, 2002: 15; Wild, 2007b: 40; Mackay & Mackay, 2010: 527 (redescription); Bezděčková, et al. 2015: 124; Feitosa, 2015c: 99.
- Senior synonym of nitidiventris: Kempf, 1961c: 201; Kempf, 1972a: 175; Bolton, 1995b: 310; Mackay & Mackay, 2010: 527.
- Distribution: Argentina, Bolivia, Brazil, Paraguay, Uruguay.
- nitidiventris. Pachycondyla striata var. nitidiventris Santschi, 1921g: 87 (w.) URUGUAY, BRAZIL (no state data).
- Type-material: 2 syntype workers.
- Type-localities: 1 worker Uruguay: Nueva Helvetia (von Steiger), 1 worker Brazil: “Bolivian frontier” (Reichensperger).
- [Note: Mackay & Mackay, 2010: 527, report 4 syntypes (“cotypes”) in NHMB, but the original description specifically states one worker from each locality. Baroni Urbani, 1973b: 134, reports only a single specimen in NHMB.]
- Type-depository: NHMB.
- Subspecies of striata: Borgmeier, 1923: 103.
- Junior synonym of striata: Kempf, 1961c: 201; Kempf, 1972a: 175; Bolton, 1995b: 307; Mackay & Mackay, 2010: 527.
Type Material
Brasil, Rio de Janeiro; Uruguay, Nueva Helvetia; Brasil, Mato Grosso, frontier with Bolivia. 2 syntype workers seen, The Natural History Museum; 4 cotypes seen, Naturhistorisches Museum, Basel. (Mackay and Mackay 2010)
The following notes on F. Smith type specimens have been provided by Barry Bolton (details):
Pachycondyla striata
Five worker syntypes in The Natural History Museum and one worker syntype in Oxford University Museum of Natural History. Labelled “Rio. 57/63.” Acc. Reg.: “1857 no. 63 (June 22). Province of Rio. Presented by Rev. Hamlet Clark.”
Three more workers in The Natural History Museum are mounted in the same way as the syntypes but have passed through the Farren White collection and had their data labels altered to read “Rio. H. Clark.” These are probably also syntypes.
Unless otherwise noted the text for the remainder of this section is reported from the publication that includes the original description.
Description
Worker
From Mackay and Mackay (2010): The worker is a relatively large (total length about 13 mm) dark brown specimen with lighter brown appendages. The mandibles have about 12 teeth. The anterior medial border of the clypeus is slightly concave, the eyes are moderately large (0.58 mm maximum diameter) located approximately one diameter from the anterior edge of the head (side view). The malar carina is absent. The scape extends past the posterior lateral corner of the head. The posterior margin of the head is concave. The pronotal carina is well developed and sharp and slightly overhangs the side of the pronotum (similar to that of Neoponera villosa). The metanotal suture is not marked on the dorsum of the mesosoma. The dorsal face of the propodeum slopes broadly into the posterior face, the propodeal spiracle is slit-shaped. The petiole is thick when viewed in profile and rectangular-shaped. The anterior face of the postpetiole is vertical and sharply bends into the dorsal face, nearly forming an angle. The dorsal face of the pygidium is concave, as in Pachycondyla impressa.
Erect hairs are present on most surfaces, including the clypeus, dorsal and ventral surfaces of the head, sides of the head, scapes, dorsum of the mesosoma, legs, including the tibiae, dorsal surface of the petiole and all surfaces of the gaster; appressed golden pubescence is sparse on most surfaces except the gaster, where it is moderately dense.
The dorsum of the head is mostly finely striate longitudinally; the striae are poorly developed on the sides of the head, but striae are present on the side of the mesosoma as well as the dorsum. The side of the petiole is nearly without striae; the dorsum of the petiole has mostly transverse very fine striae. The gaster is mostly punctate.
Queen
From Mackay and Mackay (2010): The female is similar to the worker in lacking the malar carina, having relatively large eyes, a well-developed carina on the pronotal shoulder and a similar shaped petiole. It is a large (total length 15 mm) black specimen with brown legs.
Erect and suberect golden hairs are abundant on the mandibles, clypeus, dorsal and ventral surfaces of the head, posterior margin, sides of the head, dorsum of the mesosoma, dorsum of the petiole, subpetiolar process and all surfaces of the gaster. The hairs on the legs are similar and abundant but are mostly suberect. Appressed golden pubescence is dense on most surfaces, but is fine and hides little of the surface sculpturing.
The striae are also present on the dorsum of the head but are poorly developed, especially on the sides of the head. The mandibles are mostly covered with fine striae. The dorsum of the pronotum has numerous mostly longitudinal striae, the scutum and scutellum are covered with coarse longitudinal striae. The mesopleuron and metapleuron are covered with fine nearly horizontal striae; the remainder of the ant is dull and punctate.
Male
From Mackay and Mackay (2010): The male is a large (total length 13 mm) dark brown specimen. The anterior edge of the clypeus is convex to slightly concave and the clypeus is swollen in the middle. The eyes are large and occupy approximately one half of the side of the head. The ocelli are small (maximum diameter of the median ocellus 0.12 mm), the median ocellus is located 0.2 mm from the lateral ocellus, which is greater than the diameter as the median ocellus.
The pronotal shoulder is swollen and forms a raised area, which is broadly rounded. The propodeal spiracle is slit-shaped. The petiole is thick when viewed in profile with the two faces converging dorsally and forming a rounded apex. The subpetiolar process is angulate anteriorly and gradually diminishes posteriorly. Coarse erect hairs are extremely abundant on all surfaces, especially the head, the mesosoma and petiole. Fine appressed golden pubescence is also abundant on most surfaces.
The head of the male is punctate or coriaceous, as is the mesosoma and the petiole, the gaster is finely punctate, most surfaces are dull, the mesosoma is weakly shining, as is the petiole, the gaster slightly more shining.
Karyotype
- See additional details at the Ant Chromosome Database.
- Explore: Show all Karyotype data or Search these data. See also a list of all data tables or learn how data is managed.
- n = 52, 2n = 104, karyotype = 4M+100A (Brazil) (Mariano et al., 2007; Mariano et al., 2015).
Etymology
The name of this species is derived from the Latin word stria, meaning channel or furrow, referring to the striae on numerous parts of the body, especially the pronotum. (Mackay and Mackay 2010)
References
- da Silva-Melo, A. & Giannotti, E. 2012. Division of Labor in Pachycondyla striata Fr. Smith, 1858 (Hymenoptera: Formicidae: Ponerinae). Psyche Volume 2012, Article ID 153862, 7 pages (doi:10.1155/2012/153862).
- Albuquerque, E., Prado, L., Andrade-Silva, J., Siqueira, E., Sampaio, K., Alves, D., Brandão, C., Andrade, P., Feitosa, R., Koch, E., Delabie, J., Fernandes, I., Baccaro, F., Souza, J., Almeida, R., Silva, R. 2021. Ants of the State of Pará, Brazil: a historical and comprehensive dataset of a key biodiversity hotspot in the Amazon Basin. Zootaxa 5001, 1–83 (doi:10.11646/zootaxa.5001.1.1).
- Apolinário, L.da C.M.H., Almeida, Â.A.de, Queiroz, J.M., Vargas, A.B., Almeida, F.S. 2019. Diversity and guilds of ants in different land-use Systems in Rio de Janeiro State, Brazil. Floresta e Ambiente 26, e20171152 (doi:10.1590/2179-8087.115217).
- Brown, W. L., Jr. 1958h. A review of the ants of New Zealand. Acta Hymenopterol. 1: 1-50 (page 20, Junior synonym of castanea)
- Calazans, E.G., Costa, F.V.da, Cristiano, M.P., Cardoso, D.C. 2020. Daily dynamics of an ant community in a mountaintop ecosystem. Environmental Entomology 49, 383–390 (doi:10.1093/ee/nvaa011).
- Camargo, K.S. de. 2011. Composicao e diversidade de "Poneromorfas" (Hymenoptera, Formicidae) em duas fitofisionomias de cerrado e padroes de distribuicao de "Poneromorfas", Pseudomyrmecinae e Cephalotini (Myrmicinae) para o Brasil. Thesis, Universidade de Brasilia.
- Cantone S. 2017. Winged Ants, The Male, Dichotomous key to genera of winged male ants in the World, Behavioral ecology of mating flight (self-published).
- Cantone, S., Von Zuben, C.J. 2019. The hindwings of ants: A phylogenetic analysis. Psyche: A Journal of Entomology 2019, 1–11 (doi:10.1155/2019/7929717).
- Castro-Souza, R., Pellegrini, T., Souza-Silva, M., Ferreira, R. 2019. Drivers of ant composition, richness, and trophic guilds in Neotropical iron ore cavities. International Journal of Speleology 48, 279–293 (doi:10.5038/1827-806x.48.3.2270).
- Conceição-Neto, R., França, E.C.B., Feitosa, R.M., Queiroz, J.M. 2021. Revisiting the ideas of trees as templates and the competition paradigm in pairwise analyses of ground-dwelling ant species occurrences in a tropical forest. Revista Brasileira de Entomologia 65, e20200026 (doi:10.1590/1806-9665-rbent-2020-0026).
- da Silva-Melo, A. & Giannotti, E. 2010. Nest architecture of Pachycondyla striata Fr. Smith, 1858 (Formicidae, Ponerinae). Insectes Sociaux 57, 17–22 (DOI 10.1007/s00040-009-0043-z).
- da Silva-Melo, A. & Giannotti, E. 2011. Life Expectancy and Entropy Values for Workers of Pachycondyla striata (Hymenoptera: Formicidae: Ponerinae). Sociobiology, 58, 143-150.
- Donisthorpe, H. 1949b [1948]. A fifth instalment of the Ross Collection of ants from New Guinea. Ann. Mag. Nat. Hist. 12(1): 487-506 (page 489, worker described)
- Donisthorpe, H. 1949c [1948]. A sixth instalment of the Ross Collection of ants from New Guinea. Ann. Mag. Nat. Hist. 12(1): 744-759 (page 745, Combination in pachycondyla (Bothroponera))
- Eidmann, H. 1936. Ökologisch-faunistische Studien an südbrasilianischen Ameisen. Arbeiten über physiologische und angewandte Entomologie aus Berlin-Dahlem 3:26-48, 81-114, + 1 plate, 5 text figures.
- Emery, C. 1894a. Die Ameisen von Rio Grande do Sul von Dr. H. von Jhering. Berliner Entomologische Zeitschrift 39:321-446.
- Esteves, F.A., Fisher, B.L. 2021. Corrieopone nouragues gen. nov., sp. nov., a new Ponerinae from French Guiana (Hymenoptera, Formicidae). ZooKeys 1074, 83–173 (doi:10.3897/zookeys.1074.75551).
- Forster-Figueira, C. and M. Camargo Mathias. 2002. Histological, histochemical and Morphometric Studi of female corpora allata of Pachycondyla striata ants (Hymenoptera: Ponerinae). Sociobiology 39:77-87.
- Franco, W., Ladino, N., Delabie, J.H.C., Dejean, A., Orivel, J., Fichaux, M., Groc, S., Leponce, M., Feitosa, R.M. 2019. First checklist of the ants (Hymenoptera: Formicidae) of French Guiana. Zootaxa 4674, 509–543 (doi:10.11646/zootaxa.4674.5.2).
- Garcia, F.R.M., Ovruski, S.M., Suárez, L., Cancino, J., Liburd, O.E. 2020. Biological control of tephritid fruit flies in the Americas and Hawaii: A review of the use of parasitoids and predators. Insects 11, 662. (doi:10.3390/insects11100662).
- Glaser, S.M., Feitosa, R.M., Koch, A., Goß, N., do Nascimento, F.S., Grüter, C. 2021. Tandem communication improves ant foraging success in a highly competitive tropical habitat. Insectes Sociaux. (doi:10.1007/s00040-021-00810-y).
- Karavaiev, V. 1927f. Ameisen aus dem Indo-Australischen Gebiet. III. Zb. Prats Zool. Muz. 3:3-52 [= Tr. Ukr. Akad. Nauk Fiz.-Mat. Vidd. 7:3-52]. (page 5, worker described)
- Karavaiev, V. 1935a. Neue Ameisen aus dem Indo-Australischen Gebiet, nebst Revision einiger Formen. Treubia 15: 57-118 (page 70, Replacement name: striatula)
- Kempf, W. W. 1961e. As formigas do gênero Pachycondyla Fr. Smith no Brasil (Hymenoptera: Formicidae). Rev. Bras. Entomol. 10: 189-204 (page 210?, Senior synonym of nitidiventris)
- Laciny, A. 2021. Among the shapeshifters: parasite-induced morphologies in ants (Hymenoptera, Formicidae) and their relevance within the EcoEvoDevo framework. EvoDevo 12, 2 (doi:10.1186/s13227-021-00173-2).
- Ladino, N., Feitosa, R.M. 2022. Ants (Hymenoptera: Formicidae) of the Parque Estadual São Camilo, an isolated Atlantic Forest remnant in western Paraná, Brazil. ZOOLOGIA 39: e22001 (doi:10.1590/S1984-4689.v39.e22001).
- Larabee, F.J., Suarez, A.V. 2014. The evolution and functional morphology of trap-jaw ants (Hymenoptera: Formicidae). Myrmecological News 20: 25-36.
- Lutinski, J., de Filtro, M., Baucke, L., Dorneles, F., Lutinski, C., Guarda, C. 2021. Ant assemblages (Hymenoptera: Formicidae) from areas under the direct influence of two small hydropower plants in Brazil. Brazilian Journal of Environmental Sciences (Online), 1-9 (doi:10.5327/Z217694781030).
- Mackay, W.P., Mackay, E.E. 2010. The systematics and biology of the New World ants of the genus Pachycondyla (Hymenoptera: Formicidae). Edwin Mellon Press, Lewiston.
- Marcineiro, F.S.R., Lattke, J.E. 2024. Review of ant genus Pachycondyla Smith, 1858 in Brazil (Hymenoptera: Formicidae). Zootaxa 54611, 1–62 (doi:10.11646/zootaxa.5461.1.1).
- Mariano, C.S.F., Santos, I.S., Silva, J.G., Costa, M.A., Pompolo, S.G. 2015. Citogenética e evolução do cariótipo em formigas poneromorfas. In: Delabie, J.H.C., Feitosa, R.M., Serrao, J.E., Mariano, C.S.F., Majer, J.D. (eds) As formigas poneromorfas do Brasil, 1st edn. Ilhéus, Brasil, pp 102–125 (doi:10.7476/9788574554419.0010).
- Mizumoto, N., Tanaka, Y., Valentini, G., Richardson, T. O., Annagiri, S., Pratt, S. C., Shimoji, H. 2023. Functional and mechanistic diversity in ant tandem communication. IScience 26(4), 106418 (doi:10.1016/j.isci.2023.106418).
- Morgan, E., H. Jungnickel, S. Keegans, R. do Nascimento, J. Billen, B. Gobin and F. Ito. 2003. Comparative survey of abdominal gland secretions of the ant subfamily Ponerinae. Journal of Chemical Ecology 29:95-114.
- Morgan, E., R. do Nascimento, S. Keegans and J. Billen. 1999. Comparative study of mandibular gland secretions of workers of ponerine ants. Journal of Chemical Ecology 25:1395-1409.
- Ortiz, G. and M. I. Camargo-Mathias. 2003. Morphological study of the venom gland in workers of the ant Pachycondyla striata F. Smith (Hymenoptera: Formicidae). Socio-biology 42:103-116.
- Passos, L. and P. Oliveira. 2002. Ants affect the distribution and performance of seedlings of Clusia criuva, a primarily bird-dispersed rain forest tree. Journal of Ecology 90:517-528.
- Passos, L. and S. Ferreira. 1996. Ant dispersal of Croton priscus (Euphorbiaceae) seeds in a tropical semideciduous forest in south-eastern Brazil. Biotropica 28:697-700.
- Pizo, M. and P. Oliveira. 2000. The use of fruits and seeds by ants in the Atlantic forest of southeast Brazil. Biotropica 32:81-861.
- Radchenko, A.G., Fisher, B.L., Esteves, F.A., Martynova, E.V., Bazhenova, T.N., Lasarenko, S.N. 2023. Ant type specimens (Hymenoptera, Formicidae) in the collection of Volodymyr Opanasovych Karawajew. Communication 1. Dorylinae, Poneromorpha and Pseudomyrmecinae. Zootaxa, 5244(1), 1–32 (doi:10.11646/zootaxa.5244.1.1).
- Reichensperger, A. 1936. Beltrag zur Kenntnis der Myrmecophilen und Termitophilenfauna Brasiliens und Costa Ricas. IV. (Col. Hist. Staphyl. Pselaph.). Revista de Entomologia 6:222-242.
- Rodrigues, M.S., Vilela, E.F., Azevedo, D.O., Hora, R.R. 2011. Multiple queens in founding colonies of the neotropical ant Pachycondyla striata Smith (Formicidae: Ponerinae). Neotropical Entomology 40, 293–299 (doi:10.1590/s1519-566x2011000300001).
- Rodrigues, ms, EF Vilela, DO Azevedo & RR Hora. 2011. Multiple Queens in Founding Colonies of the Neotropical Ant Pachycondyla striata Smith (Formicidae: Ponerinae). Neotropical Entomology, 40, 293-299.
- Roger, J. 1861a. Die Ponera-artigen Ameisen (Schluss). Berl. Entomol. Z. 5: 1-54 (page 6, queen described)
- Rosumek, F. B. 2017. Natural History of Ants: What We (do not) Know about Trophic and Temporal Niches of Neotropical Species. Sociobiology. 64:244-255. doi:10.13102/sociobiology.v64i3.1623
- Santos, P. P., P. D. Games, D. O. Azevedo, E. Barros, L. L. de Oliveira, H. J. D. Ramos, M. C. Baracat-Pereira, and J. E. Serrao. 2017. Proteomic analysis of the venom of the predatory ant Pachycondyla striata (Hymenoptera: Formicidae). Archives of Insect Biochemistry and Physiology. 96:17. doi:10.1002/arch.21424
- Santschi, F. 1921g. Ponerinae, Dorylinae et quelques autres formicides néotropiques. Bull. Soc. Vaudoise Sci. Nat. 54: 81-103 (page 86, male described)
- Shrestha B, Tanaka E, Hyun MW, Han JG, Kim CS, Jo JW, Han SK, Oh J, Sung JM, Sung GH. 2017. Mycosphere Essay 19. Cordyceps species parasitizing hymenopteran and hemipteran insects. Mycosphere 8(9): 1424–1442 (DOI 10.5943/mycosphere/8/9/8).
- Silva, J. P., Valadares, L., Vieira, M. E. L., Teseo, S., Châline, N. 2021. Tandem running by foraging Pachycondyla striata workers in field conditions vary in response to food type, food distance, and environmental conditions. Current Zoology 67(5), 541–549 (doi:10.1093/cz/zoab050).
- Silva-Melo, A. da; Gianotti, E. 2010. Nest architecture of Pachycondyla striata Fr. Smith, 1858 (Formicidae, Ponerinae). Insectes Sociaux 57:17-22.
- Smith, F. 1858b. Catalogue of hymenopterous insects in the collection of the British Museum. Part VI. Formicidae. London: British Museum, 216 pp. (page 106, worker described)
- Stitz, H. 1911a. Australische Ameisen. (Neu-Guinea und Salomons-Inseln, Festland, Neu-Seeland). Sitzungsber. Ges. Naturforsch. Freunde Berl. 1911: 351-381 (page 356, worker described [Unresolved junior secondary homonym of striata Smith])
- Strelnikov, T.D. 1928. Mermis parasitism in Pachycondyla striata. Psyche 35: 199-200 (doi:10.1155/1928/43012).
- Tomotake, M.E., M. Mathias, A. Yabuki and F. Caetano. 1992 Scanning electron microscopy of mandibular glands of workers and queens of the ants Pachycondyla striata, (Hymenoptera: Ponerinae). Journal of Advanced Zoology 13:1-6.
- Troya, A., Marcineiro, F., Lattke, J.E. & Longino, J. 2022. Igaponera curiosa, a new ponerine genus (Hymenoptera: Formicidae) from the Amazon. European Journal of Taxonomy 823: 82–101 (doi:10.5852/ejt.2022.823.1817).
- Ulysséa, M.A., Brandão, C.R.F. 2013. Ant species (Hymenoptera, Formicidae) from the seasonally dry tropical forest of northeastern Brazil: a compilation from field surveys in Bahia and literature records. Revista Brasileira de Entomologia 57, 217–224 (doi:10.1590/s0085-56262013005000002).
- Wheeler, G. C.; Wheeler, J. 1952c. The ant larvae of the subfamily Ponerinae - Part II. Am. Midl. Nat. 48: 604-672 (page 616, larva described)
- Wilson, E. O. 1958g. Studies on the ant fauna of Melanesia III. Rhytidoponera in western Melanesia and the Moluccas. IV. The tribe Ponerini. Bulletin of the Museum of Comparative Zoology 119: 303-371 (page 365, Replacement name: Ectomomyrmex scobinus)
- Yamane, S., Tanaka, H.O., Hasimoto, Y., Ohashi, M., Meleng, P., Itioka, T. 2021. A list of ants from Lambir Hills National Park and its vicinity, with their biological information: Part II. Subfamilies Leptanillinae, Proceratiinae, Amblyoponinae, Ponerinae, Dorylinae, Dolichoderinae, Ectatomminae and Formicinae. Contributions from the Biological Laboratory, Kyoto University 31, 87–157.
References based on Global Ant Biodiversity Informatics
- Araujo Castilho G., F. Barbosa Noll, E. R. da Silva, and E. F. dos Santos. 2011. Diversidade de Formicidae (Hymenoptera) em um fragmento de Floresta Estacional Semidecídua no Noroeste do estado de São Paulo, Brasil. R. bras. Bioci., Porto Alegre 9(2): 224-230.
- Biagiotti G., J. A. Alves Pereira, C. Rodrigues Ribas, V. Korasaki, R. Zanetti, A. C. Medeiros de Queiroz. 2013. Richness and species composition of ants in the recovery process of a Gully erosion. Cerne, Lavras 19(4): 661-668.
- Bieber A. G. D., P. D. Silva, and P. S. Oliveira. 2013. Attractiveness of Fallen Fleshy Fruits to Ants Depends on Previous Handling by Frugivores. Écoscience 20: 85-89.
- Borgmeier T. 1923. Catalogo systematico e synonymico das formigas do Brasil. 1 parte. Subfam. Dorylinae, Cerapachyinae, Ponerinae, Dolichoderinae. Archivos do Museu Nacional (Rio de Janeiro) 24: 33-103.
- Bruch C. 1914. Catálogo sistemático de los formícidos argentinos. Revista del Museo de La Plata 19: 211-234.
- Byk J., K. Del-Claro. 2010. Nectar- and pollen-gathering Cephalotes ants provide no protection against herbivory: a new manipulative experiment to test ant protective capabilities. Acta Ethol. 13: 33-38.
- Caldart V. M., S. Iop, J. A. Lutinski, and F. R. Mello Garcia. 2012. Ants diversity (Hymenoptera, Formicidae) of the urban perimeter of Chapecó county, Santa Catarina, Brazil. Revista Brasileira de Zoociências 14 (1, 2, 3): 81-94.
- Calixto J. M. 2013. Lista preliminar das especies de formigas (Hymenoptera: Formicidae) do estado do Parana, Brasil. Universidad Federal do Parana 34 pages.
- Campos R. B. F., J. H. Schoereder, and C. F. Sperber. 2007. Small-scale patch dynamics after disturbance in litter ant communities. Basic and Applied Ecology 8: 3643.
- Christianini A. V., A. J. Mayhé-Nunes, and P. S. Oliveira. 2012. Exploitation of Fallen Diaspores By Ants: Are There Ant-Plant Partner Choices? Biotropica 44: 360-367.
- Christianini A. V., and P. S. Oliveira. 2009. The relevance of ants as seed rescuers of a primarily bird-dispersed tree in the neotropical cerrado savanna. Oecologia 160: 735745.
- Christianini A. V., and P. S. Oliveira. 2013. Edge effects decrease ant-derived benefits to seedlings in a neotropical savanna. Arthropod-Plant Interactions 7: 191-199.
- Clemes Cardoso D., and J. H. Schoereder. 2014. Biotic and abiotic factors shaping ant (Hymenoptera: Formicidae) assemblages in Brazilian coastal sand dunes: the case of restinga in Santa Catarina. Florida Entomologist 97(4): 1443-1450.
- Clemes Cardoso D., and M. Passos Cristiano. 2010. Myrmecofauna of the Southern Catarinense Restinga sandy coastal plain: new records of species occurrence for the state of Santa Catarina and Brazil. Sociobiology 55(1b): 229-239.
- Cuezzo, F. 1998. Formicidae. Chapter 42 in Morrone J.J., and S. Coscaron (dirs) Biodiversidad de artropodos argentinos: una perspectiva biotaxonomica Ediciones Sur, La Plata. Pages 452-462.
- Dias N. D. S., R. Zanetti, M. S. Santos, M. F. Gomes, V. Peñaflor, S. M. F. Broglio, and J. H. C. Delabie. 2012. The impact of coffee and pasture agriculture on predatory and omnivorous leaf-litter ants. Journal of Insect Science 13:29. Available online: http://www.insectscience.org/13.29
- Dias N. S., R. Zanetti, M. S. Santos, J. Louzada, and J. H. C. Delabie. 2008. Interaction between forest fragments and adjacent coffee and pasture agroecosystems: responses of the ant communities (Hymenoptera, Formicidae). Iheringia, Sér. Zool., Porto Alegre, 98(1): 136-142.
- Diehl-Fleig E. 2014. Termites and Ants from Rio Grande do Sul, Brazil. Sociobiology (in Press).
- Drose W., L. R. Podgaiski, C. Fagundes Dias, M. de Souza Mendonca. 2019. Local and regional drivers of ant communities in forest-grassland ecotones in South Brazil: A taxonomic and phylogenetic approach. Plos ONE 14(4): e0215310.
- Emery C. 1906. Studi sulle formiche della fauna neotropica. XXVI. Bullettino della Società Entomologica Italiana 37: 107-194.
- Emery C. 1911. Hymenoptera. Fam. Formicidae. Subfam. Ponerinae. Genera Insectorum 118: 1-125.
- Favretto M. A., E. Bortolon dos Santos, and C. J. Geuster. 2013. Entomofauna from West of Santa Catarina State, South of Brazil. EntomoBrasilis 6 (1): 42-63.
- Feitosa R. dos S. M. and A. S. Ribeiro. 2005. Mirmecofauna (Hymenoptera, Formicidae) de serapilheira de uma área de Floresta Atlântica no Parque Estadual daCantareira São Paulo, Brasil. Biotemas 18: 51-71.
- Fernandes I., and J. de Souza. 2018. Dataset of long-term monitoring of ground-dwelling ants (Hymenoptera: Formicidae) in the influence areas of a hydroelectric power plant on the Madeira River in the Amazon Basin. Biodiversity Data Journal 6: e24375.
- Fernández F., and T. M. Arias-Penna. 2008. Las hormigas cazadoras en la región Neotropical. Pp. 3-39 in: Jiménez, E.; Fernández, F.; Arias, T.M.; Lozano-Zambrano, F. H. (eds.) 2008. Sistemática, biogeografía y conservación de las hormigas cazadoras de Colombia. Bogotá: Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, xiv + 609 pp.
- Fernández, F. and S. Sendoya. 2004. Lista de las hormigas neotropicales. Biota Colombiana Volume 5, Number 1.
- Figueiredo C. J. de, R. R. da Silva, C. de Bortoli Munhae, and M. S. de Castro Morini. 2013. Ant fauna (Hymenoptera: Formicidae) attracted to underground traps in Atlantic Forest. Biota Neotrop 13(1): 176-182
- Fleck M. D., E. Bisognin Cantarelli, and F. Granzotto. 2015. Register of new species of ants (Hymenoptera: Formicidae) in Rio Grande do Sul state. Ciencia Florestal, Santa Maria 25(2): 491-499.
- Forel A. 1906. Fourmis néotropiques nouvelles ou peu connues. Annales de la Société Entomologique de Belgique 50: 225-249.
- Forel A. 1907. Formiciden aus dem Naturhistorischen Museum in Hamburg. II. Teil. Neueingänge seit 1900. Mitt. Naturhist. Mus. Hambg. 24: 1-20.
- Forel A. 1907. Formicides du Musée National Hongrois. Ann. Hist.-Nat. Mus. Natl. Hung. 5: 1-42.
- Forel A. 1908. Ameisen aus Sao Paulo (Brasilien), Paraguay etc. gesammelt von Prof. Herm. v. Ihering, Dr. Lutz, Dr. Fiebrig, etc. Verhandlungen der Kaiserlich-Königlichen Zoologisch-Botanischen Gesellschaft in Wien 58: 340-418.
- Franco W., N. Ladino, J. H. C. Delabie, A. Dejean, J. Orivel, M. Fichaux, S. Groc, M. Leponce, and R. M. Feitosa. 2019. First checklist of the ants (Hymenoptera: Formicidae) of French Guiana. Zootaxa 4674(5): 509-543.
- Gomes D. S., F. S. Almeida, A. B. Vargas, and J. M. Queiroz. 2013. Resposta da assembleia de formigas na interface solo-serapilheira a um gradiente de alteração ambiental. Iheringia, Série Zoologia, Porto Alegre, 103(2):104-109
- Gomez V. E. S., and G. Z. González. 2007. Catalogo de Las Hormigas Presentes en El Museo de Historia Natural de la Universidad del Cauca. Popayán : 1-58.
- Groc S., J. H. C. Delabie, F. Fernandez, F. Petitclerc, B. Corbara, M. Leponce, R. Cereghino, and A. Dejean. 2017. Litter-dwelling ants as bioindicators to gauge the sustainability of small arboreal monocultures embedded in the Amazonian rainforest. Ecological Indicators 82: 43-49.
- Groc S., J. Orivel, A. Dejean, J. Martin, M. Etienne, B. Corbara, and J. H. C. Delabie. 2009. Baseline study of the leaf-litter ant fauna in a French Guianese forest. Insect Conservation and Diversity 2: 183-193.
- Ilha C., J. A. Lutinski, D. Von Muller Pereira, F. R. Mello Garcia. 2009. Riqueza de formigas (Hymenoptera: Formicidae) de Bacia da Sanga Caramuru, municipio de Chapeco-SC. Biotemas 22(4): 95-105.
- Iop S., V. M. Caldart, J. A. Lutinski, and F. R. Mello Garcia. 2009. Formigas urbanas da cidade de Xanxerê, Santa Catarina, Brasil. Biotemas 22(2): 55-64.
- Kamura C. M., M. S. C. Morini, C. J. Figueiredo, O. C. Bueno, and A. E. C. Campos-Farinha. 2007. Ant communities (Hymenoptera: Formicidae) in an urban ecosystem near the Atlantic Rainforest. Braz. J. Biol. 67(4): 635-641.
- Kamura, C.M., M.S.C. Morini, C.J. Figueiredo, O.C. Bueno, and A.E.C. Campos-Farinha. 2007. Comunidades de formigas (Hymenoptera: Formicidae) em um ecossistema urbano próximo à Mata Atlântica. Brazilian Journal of Biology 67(4): 635-641
- Kempf W. W. 1961. As formigas do gênero Pachycondyla Fr. Smith no Brasil (Hymenoptera: Formicidae). Revista Brasileira de Entomologia 10: 189-204.
- Kempf W. W., and K. Lenko. 1976. Levantamento da formicifauna no litoral norte e ilhas adjacentes do Estado de São Paulo, Brasil. I. Subfamilias Dorylinae, Ponerinae e Pseudomyrmecinae (Hym., Formicidae). Studia Entomologica 19: 45-66.
- Kempf, W.W. 1972. Catalago abreviado das formigas da regiao Neotropical (Hym. Formicidae) Studia Entomologica 15(1-4).
- Kusnezov N. 1953. La fauna mirmecológica de Bolivia. Folia Universitaria. Cochabamba 6: 211-229.
- Kusnezov N. 1956. Claves para la identificación de las hormigas de la fauna argentina. Idia 104-105: 1-56.
- Kusnezov N. 1978. Hormigas argentinas: clave para su identificación. Miscelánea. Instituto Miguel Lillo 61:1-147 + 28 pl.
- Lapola D. M., and H. G. Fowler. 2008. Questioning the implementation of habitat corridors: a case study in interior São Paulo using ants as bioindicators. Braz. J. Biol., 68(1): 11-20.
- Leal I. R. 2002. Diversidade de formigas do estado de Pernambuco. Pp 483 - 492 in: J. M. C. Silva & M. Tabarelli (eds.) Diagnóstico da biodiversidade de Pernambuco. Ed. Universitária da UFPE e Ed. Massangana, Recife.
- Leal I. R. and B. Cortes Lopes. 1992. Estrutura das comunidades de formigas (Hymenoptera: Formicidae) de solo e vegetacao no morro da lagoa de conceicao, Ilha de Santa Catarina, SC. Capa 5(1): 107-122.
- Luederwaldt H. 1918. Notas myrmecologicas. Rev. Mus. Paul. 10: 29-64.
- Lutinski J. A., B. C. Lopes, and A. B. B.de Morais. 2013. Diversidade de formigas urbanas (Hymenoptera: Formicidae) de dez cidades do sul do Brasil. Biota Neotrop. 13(3): 332-342.
- Lutinski J. A., F. R. Mello Garcia, C. J. Lutinska, and S. Iop. 2008. Ants diversity in Floresta Nacional de Chapecó in Santa Catarina State, Brazil. Ciência Rural, Santa Maria 38(7): 1810-1816.
- Mackay, W.P. and E.E. MacKay. 2010. The systematics and biology of the New World ants of the genus Pachycondyla (Hymenoptera: Formicidae). Edwin Mellen Press Lewiston, NY
- Majer J. D., and J. H. C. Delabie. 1994. Comparison of the ant communities of annually inundated and terra firme forests at Trombetas in the Brazilian Amazon. Insectes Sociaux 41: 343-359.
- Marinho C. G. S., R. Zanetti, J. H. C. Delabie, M. N. Schlindwein, and L. de S. Ramos. 2002. Ant (Hymenoptera: Formicidae) Diversity in Eucalyptus (Myrtaceae) Plantations and Cerrado Litter in Minas Gerais, Brazil. Neotropical Entomology 31(2): 187-195.
- Marques G. D. V., and K. Del-Claro. 2006. The Ant Fauna in a Cerrado area: The Influence of Vegetation Structure and Seasonality (Hymenoptera: Formicidae). Sociobiology 47(1): 1-18.
- Medeiros F. N. S., and P. S. Oliveira. 2009. Season-Dependent Foraging Patterns: Case Study of a Neotropical Forest-Dwelling Ant (Pachycondyla striata; Ponerinae). In: Food Exploitation by Social Insects: Ecological, Behavioral, and Theoretical Approaches. S. Jarau & M. Hrncir (editores). CRC Press, Taylor and Francis Group, Boca Raton, USA. pp. 81-95.
- Medeiros Macedo L. P., E. B. Filho, amd J. H. C. Delabie. 2011. Epigean ant communities in Atlantic Forest remnants of São Paulo: a comparative study using the guild concept. Revista Brasileira de Entomologia 55(1): 7578.
- Menozzi C. 1926. Neue Ameisen aus Brasilien. Zoologischer Anzeiger. 69: 68-72.
- Mentone T. O., E. A. Diniz, C. B. Munhae, O. C. Bueno, and M. S. C. Morini. 2011. Composition of ant fauna (Hymenoptera: Formicidae) at litter in areas of semi-deciduous forest and Eucalyptus spp., in Southeastern Brazil. Biota Neotrop. 11(2): http://www.biotaneotropica.org.br/v11n2/en/abstract?inventory+bn00511022011.
- Morini M. S. de C., C. de B. Munhae, R. Leung, D. F. Candiani, and J. C. Voltolini. 2007. Comunidades de formigas (Hymenoptera, Formicidae) em fragmentos de Mata Atlântica situados em áreas urbanizadas. Iheringia, Sér. Zool., Porto Alegre, 97(3): 246-252.
- Munhae C. B., Z. A. F. N. Bueno, M. S. C. Morini, and R. R. Silva. 2009. Composition of the Ant Fauna (Hymenoptera: Formicidae) in Public Squares in Southern Brazil. Sociobiology 53(2B): 455-472.
- Nascimento Santos M., J. H. C. Delabie, and J. M. Queiroz. 2019. Biodiversity conservation in urban parks: a study of ground-dwelling ants (Hymenoptera: Formicidae) in Rio de Janeiro City. Urban Ecosystems https://doi.org/10.1007/s11252-019-00872-8
- Oliveira Mentone T. de, E. A. Diniz, C. de Bortoli Munhae, O. Correa Bueno and M. S. de Castro Morini. 2012. Composition of ant fauna (Hymenoptera: Formicidae) at litter in areas of semi-deciduous forest and Eucalyptus spp., in Southeastern Brazil. Biota Neotrop 11(2): 237-246.
- Oliveira-Santos L. G. R., R. D. Loyola, A. B. Vargas. 2009. Canopy Traps: a Technique for Sampling Arboreal Ants in Forest Vertical Strata. Neotropical Entomology 38(5):691-694.
- Osorio Rosado J. L, M. G. de Goncalves, W. Drose, E. J. Ely e Silva, R. F. Kruger, and A. Enimar Loeck. 2013. Effect of climatic variables and vine crops on the epigeic ant fauna (Hymenoptera: Formicidae) in the Campanha region, state of Rio Grande do Sul, Brazil. J Insect Conserv 17: 1113-1123.
- Pacheco, R., R.R. Silva, M.S. de C. Morini, C.R.F. Brandao. 2009. A Comparison of the Leaf-Litter Ant Fauna in a Secondary Atlantic Forest with an Adjacent Pine Plantation in Southeastern Brazil. Neotropical Entomology 38(1):055-065
- Passos L., and P. S. Oliveira. 2002. Ants affect the distribution and performance of Clusia criuva seedlings, a primarily bird-dispersed rainforest tree. Journal of Ecology 90: 517-528.
- Passos, L. and P.S. Oliveira. 2002. Ants Affect the Distribution and Performance of Seedlings of Clusia criuva, a Primarily Bird-Dispersed Rain Forest Tree. Journal of Ecology 90(3):517-528.
- Passos, L. and P.S. Oliveira. 2003. Interactions between ants, fruits and seeds in a restinga forest in south-eastern Brazil. Journal of Tropical Ecology 19(3):261-270.
- Passos, L. and P.S. Oliviera. 2004. Interaction between Ants and Fruits of Guapira opposita (Nyctaginaceae) in a Brazilian Sandy Plain Rainforest: Ant Effects on Seeds and Seedling. Oecologia 139(3):376-382
- Pizo M. A., P. R. Guimarães, Jr., and P. S. Oliveira. 2005. Seed removal by ants from faeces produced by different vertebrate species. Ecoscience 12: 136-140.
- Pizo M. A., and P. S. Oliveira. 2000. The Use of Fruits and Seeds by Ants in the Atlantic Forest of Southeast Brazil. Biotropica 32(b): 851-861.
- Pizo M. A., and P. S. Oliveira. 2001. Size and lipid content of nonmyrmecochorous diaspores: effects on the interaction with litter-foraging ants in the Atlantic rain forest of Brazil. Plant Ecology 157: 37-52.
- Pizo, M.A. and P.S. Oliviera. 1998. Interaction between ants and seeds of a nonmyrmecochorous neotropical tree, Cabralea canjerana (Meliaceae) in the Atlantic Forest of Southeast Brazil. American Journal of Botany 85(5):669-674.
- Raimundo R. L. G., A. V. L. Freitas, and P. S. Oliveira. 2009. Seasonal patterns in activity rhythm and foraging ecology in the Neotropical forest-dwelling ant, Odontomachus chelifer (Formicidae: Ponerinae). Annals of the Entomological Society of America 102: 1151-1157.
- Raimundo, R.L.G., A.V.L. Freitas and P.S. Oliviera.2009. Seasonal Patterns in Activity Rhythm and Foraging Ecology in the Neotropical Forest-Dwelling Ant, Odontomachus chelifer (Formicidae: Ponerinae). Ann. Entomol. Soc. Am. 102(6): 1151-1157
- Ramos L. S., R. Z. B. Filho, J. H. C. Delabie, S. Lacau, M. F. S. dos Santos, I. C. do Nascimento, and C. G. S. Marinho. 2003. Ant communities (Hymenoptera: Formicidae) of the leaf-litter in cerrado stricto sensu areas in Minas Gerais, Brazil. Lundiana 4(2): 95-102.
- Resende J. J., G. M. de M. Santos, I. C. do Nascimento, J. H. C. Delabie, and E. M. da Silva. 2011. Communities of ants (Hymenoptera Formicidae) in different Atlantic rain forest phytophysionomies. Sociobiology 58(3): 779-799.
- Ribas C. R., F. A. Schmidt, R. R. C. Solar, R. B. F. Campos, C. L. Valentim, and J. H. Schoereder. 2012. Ants as Indicators of the Success of Rehabilitation Efforts in Deposits of Gold Mining Tailings. Restoration Ecology 20(6): 712722.
- Rodrigues de Souza D., E. Stingel, L. C. de Almeida, M. A. Lazarini, C. de Bortoli Munhae, A. J. Mayhe-Nunes, O. Correa Bueno, and M. Santina de C. Morini. 2010. Ant Diversity in a Sugarcane Culture without the Use of Straw Burning in Southeast, São Paulo, Brazil. American Journal of Agricultural and Biological Sciences 5 (2): 183-188.
- Rodrigues de Souza D., E. Stingel, L. C. de Almeida, M. A. Lazarini, C. de Bortoli Munhae, O. Correa Bueno, C. R. Archangelo, and M. Santina de C. Morini. 2010. Field methods for the study of ants in sugarcane plantations in Southeastern Brazil. Sci. Agric. (Piracicaba, Braz.) 67(6): 651-657.
- Rosa da Silva R. 1999. Formigas (Hymenoptera: Formicidae) do oeste de Santa Catarina: historico das coletas e lista atualizada das especies do Estado de Santa Catarina. Biotemas 12(2): 75-100.
- Rosa da Silva R., and B. Cortes Lopes. 1997. Ants (Hymenoptera: Formicidae) from Atlantic rainforest at Santa Catarina Island, Brazil: two years of sampling. Rev. Biol. Trop. 45(4): 1641-1648.
- Rosumek, F.B., M.A. Ulyssea, B.C. Lopes, J. Steiner. 2008. Formigas de solo e de bromélias em uma área de Mata Atlântica, Ilha de Santa Catarina, sul do Brasil: Levantamento de espécies e novos registros. Revista Biotemas 21(4):81-89.
- Salinas P. J. 2010. Catalogue of the ants of the Táchira State, Venezuela, with notes on their biodiversity, biogeography and ecology (Hymenoptera: Formicidae: Amblyioponinae, Ponerinae, Proceratiinae, Myrmicinae, Ecitoninae, Formicinae, Pseudomyrmecinae, Dolichoderinae). Boletín de la SEA 47: 315-328.
- Santoandre S., J. Filloy, G. A. Zurita, and M. I. Bellocq. 2019. Ant taxonomic and functional diversity show differential response to plantation age in two contrasting biomes. Forest Ecology and Management 437: 304-313.
- Santos Lopes J. F., N. Martins dos Reis Hallack, T. Archanjo de Sales, M. Silva Brugger, L. F. Ribeiro, I. N. Hastenreiter, and R. da Silva Camargo. 2012. Comparison of the Ant Assemblages in Three Phytophysionomies: Rocky Field, Secondary Forest, and Riparian ForestA Case Study in the State Park of Ibitipoca, Brazil. Psyche doi:10.1155/2012/928371
- Santos M. S., J. N. C. Louzada, N. Dias, R. Zanetti, J. H. C. Delabie, and I. C. Nascimento. 2006. Litter ants richness (Hymenoptera, Formicidae) in remnants of a semi-deciduous forest in the Atlantic rain forest, Alto do Rio Grande region, Minas Gerais, Brazil. Iheringia, Sér. Zool., Porto Alegre, 96(1): 95-101.
- Santos-Junior L. C., J. M. Saraiva, R. Silvestre, and W. F. Antonialli-Junior. 2014. Evaluation of Insects that Exploit Temporary Protein Resources Emphasizing the Action of Ants (Hymenoptera, Formicidae) in a Neotropical Semi-deciduous Forest. Sociobiology 61(1): 43-51
- Santschi F. 1916. Formicides sudaméricains nouveaux ou peu connus. Physis (Buenos Aires). 2: 365-399.
- Santschi F. 1921. Ponerinae, Dorylinae et quelques autres formicides néotropiques. Bulletin de la Société Vaudoise des Sciences Naturelles 54: 81-103.
- Santschi F. 1925. Fourmis des provinces argentines de Santa Fe, Catamarca, Santa Cruz, Córdoba et Los Andes. Comunicaciones del Museo Nacional de Historia Natural "Bernardino Rivadavia" 2: 149-168.
- Schmidt K., R. Corbetta, and A.J. Aires de Carmargo. 2005. Formigas (Hymenoptera: Formicidae) da Ilha João da Cunha, SC: composição e diversidade. Biotemas, 18 (1): 57 - 71
- Silva R. R., R. S. Machado Feitosa, and F. Eberhardt. 2007. Reduced ant diversity along a habitat regeneration gradient in the southern Brazilian Atlantic Forest. Forest Ecology and Management 240: 61-69.
- Silva R.R., and C. R. F. Brandao. 2014. Ecosystem-Wide Morphological Structure of Leaf-Litter Ant Communities along a Tropical Latitudinal Gradient. PLoSONE 9(3): e93049. doi:10.1371/journal.pone.0093049
- Silva T. S. R., and R. M. Feitosa. 2019. Using controlled vocabularies in anatomical terminology: A case study with Strumigenys (Hymenoptera: Formicidae). Arthropod Structure and Development 52: 1-26.
- Silvestre R., C. R. F. Brandão, and R. R. Silva da 2003. Grupos funcionales de hormigas: el caso de los gremios del cerrado. Pp. 113-148 in: Fernández, F. (ed.) 2003. Introducción a las hormigas de la región Neotropical. Bogotá: Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, xxvi + 424 pp.
- Soares S. A., D. Lange, and W. F. Antoniali Junior. 2006. Communities of Epigaeic ants (Hymenoptera: Formicidae) in an area of reforestation and in native forest. Sociobiology 49(3): 251-263.
- Soares S. A., W. F. Antoniali Junior, and S. E. Lima-Junior. 2010. Diversidade de formigas epigéicas (Hymenoptera, Formicidae) em dois ambientes no Centro-Oeste do Brasil. Revista Brasileira de Entomologia 54(1): 7681.
- Sobrinho T. G., and J. H. Schoereder. 2007. Edge and shape effects on ant (Hymenoptera: Formicidae) species richness and composition in forest fragments. Biodivers Conserv 16: 14591470.
- Sobrinho T., J. H. Schoereder, C. F. Sperber, and M. S. Madureira. 2003. Does fragmentation alter species composition in ant communities (Hymenoptera: Formicidae)? Sociobiology 42(2): 329-342.
- Suguituru S. S., D. R. de Souza, C. de Bortoli Munhae, R. Pacheco, and M. S. de Castro Morini. 2011. Diversidade e riqueza de formigas (Hymenoptera: Formicidae) em remanescentes de Mata Atlântica na Bacia Hidrográfica do Alto Tietê, SP. Biota Neotrop. 13(2): 141-152.
- Suguituru S. S., M. Santina de Castro Morini, R. M. Feitosa, and R. Rosa da Silva. 2015. Formigas do Alto Tiete. Canal 6 Editora 458 pages
- Suguituru S. S., R. Rosa Silva, D. R. de Souza, C. de Bortoli Munhae, and M. Santina de Castro Morini. Ant community richness and composition across a gradient from Eucalyptus plantations to secondary Atlantic Forest. Biota Neotrop. 11(1): 369-376.
- Ulyssea M. A., B. C. Lopes, A. Zillidens, J. Steiner. 2007. Ants associated to Nidularium innocentii and Aechmea lindenii (Bromeliaceae) in atlantic rainforest in southern Brazil. Biológico, São Paulo 69(2): 319-324.
- Ulyssea M.A., C. E. Cereto, F. B. Rosumek, R. R. Silva, and B. C. Lopes. 2011. Updated list of ant species (Hymenoptera, Formicidae) recorded in Santa Catarina State, southern Brazil, with a discussion of research advances and priorities. Revista Brasileira de Entomologia 55(4): 603-611.
- Vernalha M. M. 1952. Algumas formigas que ocorrem no estado do Parana. Arquivos de Biologia e Tecnologia 7: 43-51.
- Vittar, F. 2008. Hormigas (Hymenoptera: Formicidae) de la Mesopotamia Argentina. INSUGEO Miscelania 17(2):447-466
- Wheeler W. M. 1930. Two mermithergates of Ectatomma. Psyche (Cambridge) 37: 48-54.
- Wild A. L. 2002. The genus Pachycondyla (Hymenoptera: Formicidae) in Paraguay. Boletín del Museo Nacional de Historia Natural del Paraguay 14: 1-18
- Wild, A. L. "A catalogue of the ants of Paraguay (Hymenoptera: Formicidae)." Zootaxa 1622 (2007): 1-55.
- Zolessi L. C. de, Y. P. Abenante, and M. E. de Philippi. 1988. Lista sistematica de las especies de Formicidos del Uruguay. Comun. Zool. Mus. Hist. Nat. Montev. 11: 1-9.
- Zolessi L. C. de; Y. P. de Abenante, and M. E. Philippi. 1989. Catálogo sistemático de las especies de Formícidos del Uruguay (Hymenoptera: Formicidae). Montevideo: ORCYT Unesco, 40 + ix pp.
- da Silva Araujo, M., Castro Della Lucia, T.M., DA VEIGA, Clayton E y CARDOSO DO NASCIMENTO, Ivan. 2004. Efeito da queima da palhada de cana-de-açúcar sobre comunidade de formicídeos. Ecol. austral. 14(2): 191-200.
- da Silva R. R., and R. Silvestre. 2000. Diversidade de formigas (Hymenoptera: Formicidae) em Seara, oeste de Santa Catarina. Biotemas 13(2): 85-105.
- da Silva, R.R. and R. Silvestre. 2004. Riqueza da fauna de formigas (Hymenoptera: Formicidae) que habita as camadas superficiais do solo em Seara, Santa Catarina. Papéis Avulsos de Zoologia (São Paulo) 44(1): 1-11
- da Silva, R.R., C.R.F. Brandao, and R. Silvestre. 2004. Similarity Between Cerrado Localities in Central and Southeastern Brazil Based on the Dry Season Bait Visitors Ant Fauna. Studies on Neotropical Fauna and Environment 39(3):191-199.
- da Silva-Melo, A. and E. Giannotti. 2010. Nest architecture of Pachycondyla striata Fr. Smith, 1858 (Formicidae, Ponerinae). Insectes Sociaux 57(1):17-22.
- de Souza D. R., S. G. dos Santos, C. de B. Munhae, and M. S. de C. Morini. 2012. Diversity of Epigeal Ants (Hymenoptera: Formicidae) in Urban Areas of Alto Tietê. Sociobiology 59(3): 703-117.
- de Zolessi, L.C., Y.P. de Abenante and M.E. Philippi. 1987. Lista sistemática de las especies de formícidos del Uruguay. Comunicaciones Zoologicas del Museo de Historia Natural de Montevideo 11(165):1-9
- de Zolessi, L.C., Y.P. de Abenante and M.E. Phillipi. 1989. Catalago Systematico de las Especies de Formicidos del Uruguay (Hymenoptera: Formicidae). Oficina Regional de Ciencia y Technologia de la Unesco para America Latina y el Caribe- ORCYT. Montevideo, Uruguay
- dos Santos Pereira, M.P., J.M. Queiroz, R. Valcarcel & A.J. Mayhé-Nunes. 2007. FAUNA DE FORMIGAS COMO FERRAMENTA PARA MONITORAMENTO DE ÁREA DEMINERAÇÃO REABILITADA NA ILHA DA MADEIRA, ITAGUAÍ, RJ. Ciência Florestal, Vol. 17, No. 3, 2007, pp. 197-204.
- Pages using DynamicPageList3 parser function
- North subtropical
- Tropical
- South subtropical
- South temperate
- Phorid fly Associate
- Host of Apocephalus piliventris
- Nematode Associate
- Host of Mermithidae (Mermithidae (unspecified "Mermis")
- Fungus Associate
- Host of Ophiocordyceps australis
- Karyotype
- Species
- Extant species
- Formicidae
- Ponerinae
- Ponerini
- Pachycondyla
- Pachycondyla striata
- Ponerinae species
- Ponerini species
- Pachycondyla species
- Ssr