Odontomachus chelifer
| Odontomachus chelifer | |
|---|---|

| |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Arthropoda |
| Class: | Insecta |
| Order: | Hymenoptera |
| Family: | Formicidae |
| Subfamily: | Ponerinae |
| Tribe: | Ponerini |
| Genus: | Odontomachus |
| Species: | O. chelifer |
| Binomial name | |
| Odontomachus chelifer (Latreille, 1802)
| |
| Synonyms | |
| |


{kind=link}
De la Mora et al. (2016) found this species nesting in logs found in coffee farms and forests in the Soconusco region of Chiapas, Mexico. A small percentage of pupae were found to be parasitized by eucharitid wasps.
| At a Glance | • Polygynous |
Identification
A member of the Odontomachus haematodus group.
Keys including this Species
Distribution
Latitudinal Distribution Pattern
Latitudinal Range: 27.158° to -64.36°.
| North Temperate |
North Subtropical |
Tropical | South Subtropical |
South Temperate |
- Source: AntMaps
Distribution based on Regional Taxon Lists
Neotropical Region: Argentina, Bolivia, Brazil, Colombia, Costa Rica, French Guiana, Guatemala, Guyana, Honduras, Mexico, Panama, Paraguay, Peru, Suriname, Venezuela.
Distribution based on AntMaps
Distribution based on AntWeb specimens
Check data from AntWeb
Countries Occupied
| Number of countries occupied by this species based on AntWiki Regional Taxon Lists. In general, fewer countries occupied indicates a narrower range, while more countries indicates a more widespread species. |

|
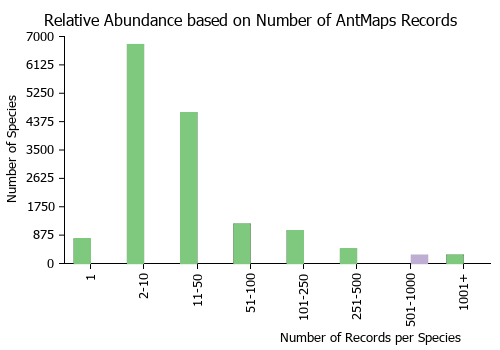
Estimated Abundance
| Relative abundance based on number of AntMaps records per species (this species within the purple bar). Fewer records (to the left) indicates a less abundant/encountered species while more records (to the right) indicates more abundant/encountered species. |
|
Biology
Rosumek (2017) examined trophic and temporal niches of ants in a secondary Atlantic forest (Florianópolis, south Brazil 27°31′38″S 48°30′15″W / 27.52722°S 48.50417°W, between December 2015 and January 2016. For this species: Odontomachus chelifer and Pachycondyla striata are two widespread species that differ radically in morphology but are similar in many aspects. It is appropriate to discuss both together. They are one of the most conspicuous elements of the southern Atlantic forest ground fauna, due to their abundance, solitary foraging mode and large size. Several accounts show these species occupy a multitude of functional roles and have a broad trophic niche. Observation of nest entrances showed that 80-90% of the items carried by P. striata were arthropod parts, mostly termites and other ants, with the remainder consisting of plant material (Giannotti & Machado, 1991; Medeiros & Oliveira, 2009). Through direct observation, Medeiros and Oliveira (2009) also showed that scavenging accounts for more than 80% of its foraging behavior. On the other hand, Fowler (1980) reported O. chelifer prefers certain termite species in the laboratory, and qualitatively stated that in the field prey consisted almost entirely of termites. In the Atlantic forest, all items carried to nests were arthropods, mainly termites, but other animal groups accounted for 60% of them (Raimundo et al., 2009). Scavenging was also cited in this study, although not quantified. Both O. chelifer and P. striata were recorded on experimental carcasses predating the larvae and pupae of necrophagous insects (Paula et al., 2016).
Other important items used by the two species are seeds with elaiosomes and other fallen diaspores rich in proteins and lipids, frequently collected from the ground in the Atlantic forest (Pizo & Oliveira, 2000; Passos & Oliveira, 2002, 2004). Field records on use of liquid sugars are scant and qualitative. Odontomachus chelifer was not observed using extra-floral nectaries by Raimundo et al. (2009), while there is one record for P. striata (in Cerrado, the Brazilian savannah – Byk & Del-Claro, 2010).
The results presented here are mostly consistent with this broader picture. Both species more frequently foraged upon dead insects, sucrose and large prey. In fact, they were the only two species consistently recorded on crickets. The low frequency in termite baits is unexpected and probably represents a methodological artifact. These large solitary foragers were observed quickly collecting termites (even glued ones) and leaving the baits in a few minutes, contrary to ants of smaller species that recruited to them. In cricket baits, however, the two species spent more time trying to carry out the tied cricket or dismembering it. Smaller ants frequently took advantage of this to grab the remains or lick spilled hemolymph. This largely contributed to the richness found in this bait and could happen in nature, whenever predators kill prey too large to carry them out at once.
Besides predation, scavenging was a common behavior, and P. striata in particular would prioritize it whenever possible, in accordance with what was observed by Medeiros and Oliveira (2009). This could be result of their morphology, because the triangular mandibles may be more suited to chop large carcasses than the trap-jaws of O. chelifer. In addition, it could be an effect of competition. The two species were never found at the same bait, and co-occurred in just 10 of the 62 points where they were recorded. The two were previously reported to avoid each other, but, when agonistic interactions occur, P. striata usually is the winner, and can steal the food or kill (and eat) O. chelifer (Medeiros & Oliveira, 2009; Raimundo et al., 2009). Thus, P. striata could displace O. chelifer and maintain control of a valuable resource such as dead arthropods through tandem recruitment (Medeiros & Oliveira, 2009; Silva-Melo & Giannotti, 2012), while cooperative foraging behavior was not observed in O. chelifer (Raimundo et al., 2009). Effectively, the average numbers of workers per bait was smaller for the latter species (O. chelifer = 1.4 ± 0.9; P. striata = 2.5 ± 2; Mann-Whitney, z = -2.27, p = 0.02).
Both species used sucrose frequently and rarely foraged on melezitose (anolog for honeydew). They were never observed foraging on trees or low vegetation in this study, which fits previous accounts (Fowler, 1980; Medeiros & Oliveira 2009), so it is unlikely that they commonly use nectar as food source. This behavior also should limit honeydew use by them.
Past results have shown that these species show a preference for day (P. striata) or night foraging (O. chelifer). No preference in their time of foraging was found in this study.
Lange et al. (2015) - This species was found in semideciduous mesophytic forest pitfall-trap samples. The study examined the ant community within a patch (11.2 hectares) of forest in an urban park (Araguari, Brazil).
Association with Other Organisms
 Explore: Show all Associate data or Search these data. See also a list of all data tables or learn how data is managed.
Explore: Show all Associate data or Search these data. See also a list of all data tables or learn how data is managed.
- This species is a host for the eucharitid wasp Latina rugosa (a parasite) (Universal Chalcidoidea Database) (associate, primary host).
- This species is a host for the phorid fly Apocephalus catholicus (a parasite) (Brown et al., 2015) (injured).
- This species is a host for the phorid fly Apocephalus catholicus (a parasite) (phorid.net) (attacked).
- This species is a host for the phorid fly Apocephalus paldiae (a parasite) (Brown et al., 2015) (injured).
- This species is a host for the phorid fly Apocephalus paldiae (a parasite) (phorid.net) (attacked).
- This species is a host for the phorid fly Dohrniphora sp. (not D. longi-gp) (a parasite) (Brown et al., 2015) (injured).
- This species is a prey for the phorid fly Dohrniphora sp. (a predator) (Quevillon, 2018).
- This species is a host for the nematode Mermithidae (unspecified "Mermix") (a parasite) in Neotropics (Wheeler, 1928; Laciny, 2021).
Castes
Worker
 
| |
| . | |
Images from AntWeb
   
| |
| Worker. Specimen code casent0178692. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by MIZA, Maracay, Venezuela. |
Nomenclature
The following information is derived from Barry Bolton's Online Catalogue of the Ants of the World.
- chelifer. Formica chelifera Latreille, 1802c: 188, pl. 8, fig. 51 (w.) (no state data).
- Type-material: syntype workers (number not stated).
- Type-locality: unknown. Latreille states, “J’ignore sa patrie......Je ne connois point son lieu natal”.
- Type-depository: MNHN.
- Forel, 1909a: 252 (q.m.); Wheeler, G.C. & Wheeler, J. 1952c: 651 (l.).
- [Misspelled as cheliferus by Smith, F. 1858b: 77.]
- Combination in Odontomachus: Latreille, 1809: 128.
- Status as species: Leach, 1815: 147; Lepeletier de Saint-Fargeau, 1835: 186; Le Guillou, 1842: 318; Smith, F. 1858b: 77; Roger, 1861a: 27; Roger, 1863b: 22; Mayr, 1863: 436; Mayr, 1865: 63 (footnote); Forel, 1886a: 217; Emery, 1888c: 353; Emery, 1890a: 65; Emery, 1890b: 44; Emery, 1892d: 560 (in key); Dalla Torre, 1893: 49; von Jhering, 1894: 381; Emery, 1894k: 50; Forel, 1895b: 118; Emery, 1896h: 625; Forel, 1899c: 19; Emery, 1904b: 595; Forel, 1905b: 156; Emery, 1906c: 118; Forel, 1908e: 64; Forel, 1909a: 252; Emery, 1911d: 114; Forel, 1912c: 28; Forel, 1914e: 9; Bruch, 1914: 213; Wheeler, W.M. 1916c: 3; Gallardo, 1918b: 96; Luederwaldt, 1918: 36; Mann, 1922: 18; Borgmeier, 1923: 77; Wheeler, W.M. 1925a: 10; Eidmann, 1936a: 37; Kempf, 1962b: 17 (in key); Kempf, 1972a: 170; Kempf & Lenko, 1976: 59; Brown, 1976a: 103; Brandão, 1991: 364; Bolton, 1995b: 295; Wild, 2007b: 39; Rodriguez, J. 2008: 161; Branstetter & Sáenz, 2012: 262; Bezděčková, et al. 2015: 124; Feitosa, 2015c: 99; Fernández & Guerrero, 2019: 538.
- Senior synonym of emacerata: Brown, 1976a: 103; Brandão, 1991: 364; Bolton, 1995b: 295.
- Senior synonym of leptocephalus: Emery, 1911d: 114; Borgmeier, 1923: 77; Kempf, 1972a: 170; Brown, 1976a: 103; Bolton, 1995b: 295.
- Senior synonym of theresiae: Forel, 1899c: 19; Gallardo, 1918b: 96; Brown, 1976a: 103; Brandão, 1991: 364; Bolton, 1995b: 295.
- Distribution: Argentina, Bolivia, Brazil, Colombia, Costa Rica, French Guiana, Guatemala, Guyana, Honduras, Mexico, Panama, Paraguay, Peru, Suriname, Venezuela.
- emacerata. Odontomachus chelifer var. emacerata Santschi, 1922d: 243 (w.) BRAZIL (Santa Catarina), ARGENTINA (Santiago del Estero).
- Type-material: syntype workers (number not stated).
- Type-localities: Brazil: Santa Catarina, Blumenau (Reichensperger), and Argentina: Chaco de Santiago del Estero (Wagner).
- Type-depository: NHMB.
- Subspecies of chelifer: Kempf, 1972a: 170.
- Junior synonym of chelifer: Brown, 1976a: 103; Brandão, 1991: 364; Bolton, 1995b: 295.
- leptocephalus. Odontomachus chelifer var. leptocephalus Emery, 1890b: 45 (in text), pl. 5, fig. 2 (w.) BRAZIL (Rio Grande do Sul).
- Type-material: holotype worker (mermithergate).
- Type-locality: Brazil: Rio Grande do Sul (v. Jhering).
- Type-depository: MSNG.
- Subspecies of chelifer: Dalla Torre, 1893: 49; Forel, 1895b: 118.
- Junior synonym of chelifer: Emery, 1911d: 114; Borgmeier, 1923: 77; Kempf, 1972a: 170; Brown, 1976a: 103; Bolton, 1995b: 296.
- theresiae. Odontomachus haematodes r. theresiae Forel, 1895a: 41 (w.) MEXICO (Veracruz).
- Type-material: holotype(?) worker.
- [Note: no indication of number of specimens is given.]
- Type-locality: Mexico: Colorado, nr Veracruz (Pr. Thérèse de Bavière).
- Type-depository: MHNG.
- Emery, 1901a: 53 (m.).
- Subspecies of chelifer: Emery, 1901a: 53; Emery, 1911d: 114; Forel, 1912c: 28; Borgmeier, 1923: 77; Kempf, 1972a: 170.
- Junior synonym of chelifer: Forel, 1899c: 19; Gallardo, 1918b: 96; Brown, 1976a: 103; Brandão, 1991: 364; Bolton, 1995b: 297.
Description
Karyotype
- See additional details at the Ant Chromosome Database.
- Explore: Show all Karyotype data or Search these data. See also a list of all data tables or learn how data is managed.
- n = 22, 2n = 44, karyotype = 4SM+40T (4M + 40A) (Brazil) (Santos et al., 2010; Mariano et al., 2015).
References
- Aguiar, H.J.A.C., Barros, L.A.C., Silveira, L.I., Petitclerc, F., Etienne, S., Orivel, J. 2020. Cytogenetic data for sixteen ant species from North-eastern Amazonia with phylogenetic insights into three subfamilies. Comparative Cytogenetics 14(1): 43–60 (doi:10.3897/CompCytogen.v14i1.46692).
- Albrecht, M., Gotelli, N.J. 2001. Spatial and temporal niche partitioning in grassland ants. Oecologia 126, 134–141 (doi:10.1007/s004420000494).
- Albuquerque, E., Prado, L., Andrade-Silva, J., Siqueira, E., Sampaio, K., Alves, D., Brandão, C., Andrade, P., Feitosa, R., Koch, E., Delabie, J., Fernandes, I., Baccaro, F., Souza, J., Almeida, R., Silva, R. 2021. Ants of the State of Pará, Brazil: a historical and comprehensive dataset of a key biodiversity hotspot in the Amazon Basin. Zootaxa 5001, 1–83 (doi:10.11646/zootaxa.5001.1.1).
- Apolinário, L.da C.M.H., Almeida, Â.A.de, Queiroz, J.M., Vargas, A.B., Almeida, F.S. 2019. Diversity and guilds of ants in different land-use Systems in Rio de Janeiro State, Brazil. Floresta e Ambiente 26, e20171152 (doi:10.1590/2179-8087.115217).
- Bieber, A.G.D., Pereira, E.R., Bottcher, C. 2020. Flower perianth: first observations on a novel diet item for the ponerine ant Odontomachus chelifer (Latreille). Revista Brasileira de Entomologia 64: e20200019 (doi:10.1590/1806-9665-rbent-2020-0019).
- Boudinot, B.E., Richter, A.K., Hammel, J.U., Szwedo, J., Bojarski, B., Perrichot, V. 2022. Genomic-phenomic reciprocal illumination: Desyopone hereon gen. et sp. nov., an exceptional Aneuretine-like fossil ant from Ethiopian amber (Hymenoptera: Formicidae: Ponerinae). Insects 13(9), 796 (doi:10.3390/insects13090796).
- Brown, B., Kung, G.-A., Porras, W. 2015. A new type of ant-decapitation in the Phoridae (Insecta: Diptera). Biodiversity Data Journal 3, e4299 (doi:10.3897/bdj.3.e4299).
- Brown, W. L., Jr. 1976c. Contributions toward a reclassification of the Formicidae. Part VI. Ponerinae, tribe Ponerini, subtribe Odontomachiti. Section A. Introduction, subtribal characters. Genus Odontomachus. Stud. Entomol. 19: 67-171 (page 103, Senior synonym of emacerata, Senior synonym of theresiae)
- Calazans, E.G., Costa, F.V.da, Cristiano, M.P., Cardoso, D.C. 2020. Daily dynamics of an ant community in a mountaintop ecosystem. Environmental Entomology 49, 383–390 (doi:10.1093/ee/nvaa011).
- Camargo, K.S. de. 2011. Composicao e diversidade de "Poneromorfas" (Hymenoptera, Formicidae) em duas fitofisionomias de cerrado e padroes de distribuicao de "Poneromorfas", Pseudomyrmecinae e Cephalotini (Myrmicinae) para o Brasil. Thesis, Universidade de Brasilia.
- Castro-Souza, R., Pellegrini, T., Souza-Silva, M., Ferreira, R. 2019. Drivers of ant composition, richness, and trophic guilds in Neotropical iron ore cavities. International Journal of Speleology 48, 279–293 (doi:10.5038/1827-806x.48.3.2270).
- de la Mora, A., G. Perez-Lachaud, J. P. Lachaud, and S. M. Philpott. 2015. Local and Landscape Drivers of Ant Parasitism in a Coffee Landscape. Environmental Entomology. 44:939-950. doi:10.1093/ee/nvv071
- Emery, C. 1911e. Hymenoptera. Fam. Formicidae. Subfam. Ponerinae. Genera Insectorum 118: 1-125 (page 114, Senior synonym of leptocephalus)
- Forel, A. 1899b. Formicidae. [part]. Biol. Cent.-Am. Hym. 3: 1-24 (page 19, Senior synonym of theresiae)
- Forel, A. 1909a. Ameisen aus Guatemala usw., Paraguay und Argentinien (Hym.). Dtsch. Entomol. Z. 1909: 239-269 (page 252, queen, male described)
- Franco, W., Ladino, N., Delabie, J.H.C., Dejean, A., Orivel, J., Fichaux, M., Groc, S., Leponce, M., Feitosa, R.M. 2019. First checklist of the ants (Hymenoptera: Formicidae) of French Guiana. Zootaxa 4674, 509–543 (doi:10.11646/zootaxa.4674.5.2).
- Glaser, S.M., Feitosa, R.M., Koch, A., Goß, N., do Nascimento, F.S., Grüter, C. 2021. Tandem communication improves ant foraging success in a highly competitive tropical habitat. Insectes Sociaux. (doi:10.1007/s00040-021-00810-y).
- Ladino, N., Feitosa, R.M. 2022. Ants (Hymenoptera: Formicidae) of the Parque Estadual São Camilo, an isolated Atlantic Forest remnant in western Paraná, Brazil. ZOOLOGIA 39: e22001 (doi:10.1590/S1984-4689.v39.e22001).
- Lange, D., A. A. Vilela, G. Erdogmus, A. B. Barbosa, S. C. Costa, and V. Stefani. 2015. Temporal dynamic of foraging of epigeic ants in an urban forest fragment. Bioscience Journal. 31:1501-1511.
- Larabee, F.J., Suarez, A.V. 2014. The evolution and functional morphology of trap-jaw ants (Hymenoptera: Formicidae). Myrmecological News 20: 25-36.
- Latreille, P. A. 1809. Genera crustaceorum et insectorum secundum ordinem naturalem in familias disposita, iconibus exemplisque plurimus explicata. Tomus 4. Parisiis et Argentorati [= Paris and Strasbourg]: A. Koenig, 399 pp. (page 128, Combination in Odontomachus)
- Latreille, P.A. 1802. Histoire naturelle des fourmis, et recueil de mémoires et d'observations sur les abeilles, les araignées, les faucheurs, et autres insectes. Paris: Impr. Crapelet (chez T. Barrois), xvi + 445 pp.
- Lutinski, J., de Filtro, M., Baucke, L., Dorneles, F., Lutinski, C., Guarda, C. 2021. Ant assemblages (Hymenoptera: Formicidae) from areas under the direct influence of two small hydropower plants in Brazil. Brazilian Journal of Environmental Sciences (Online), 1-9 (doi:10.5327/Z217694781030).
- Mariano, C.S.F., Santos, I.S., Silva, J.G., Costa, M.A., Pompolo, S.G. 2015. Citogenética e evolução do cariótipo em formigas poneromorfas. In: Delabie, J.H.C., Feitosa, R.M., Serrao, J.E., Mariano, C.S.F., Majer, J.D. (eds) As formigas poneromorfas do Brasil, 1st edn. Ilhéus, Brasil, pp 102–125 (doi:10.7476/9788574554419.0010).
- Mendoza-Guido, B., Rodríguez-Hernández, N., Ivens, A.B.F., von Beeren, C., Murillo-Cruz, C., Zuniga-Chaves, I., Łukasik, P., Sanchez, E., Kronauer, D.J.C., Pinto-Tomás, A.A. 2023. Low diversity and host specificity in the gut microbiome community of Eciton army ants (Hymenoptera: Formicidae: Dorylinae) in a Costa Rican rainforest. Myrmecological News 33: 19-34 (doi:10.25849/MYRMECOL.NEWS_033:019).
- Meurville, M.-P., LeBoeuf, A.C. 2021. Trophallaxis: the functions and evolution of social fluid exchange in ant colonies (Hymenoptera: Formicidae). Myrmecological News 31: 1-30 (doi:10.25849/MYRMECOL.NEWS_031:001).
- Moura, M.N., Cardoso, D.C., Cristiano, M.P. 2020. The tight genome size of ants: diversity and evolution under ancestral state reconstruction and base composition. Zoological Journal of the Linnean Society, zlaa135 (doi:10.1093/zoolinnean/zlaa135).
- Paul, J. Gronenberg, W. 1999. Optimizing force and velocity: mandible muscle fibre attachments in ants. Journal of Experimental Biology 202, 797-808.
- Rodrigues, M.S., Vilela, E.F., Azevedo, D.O., Hora, R.R. 2011. Multiple queens in founding colonies of the neotropical ant Pachycondyla striata Smith (Formicidae: Ponerinae). Neotropical Entomology 40, 293–299 (doi:10.1590/s1519-566x2011000300001).
- Rosumek, F. B. 2017. Natural History of Ants: What We (do not) Know about Trophic and Temporal Niches of Neotropical Species. Sociobiology. 64:244-255. doi:10.13102/sociobiology.v64i3.1623
- Santos-Junior, L.C., Silva, E.P., Antonialli-Junior, W.F. 2021. Do Odontomachus brunneus nestmates request for help and are taken care of when caught? Sociobiology 68, e6022 (doi:10.13102/sociobiology.v68i3.6022).
- Tibcherani, M., Aranda, R., Mello, R.L. 2020. Time to go home: The temporal threshold in the regeneration of the ant community in the Brazilian savanna. Applied Soil Ecology 150, 103451 (doi:10.1016/j.apsoil.2019.103451).
- Ulysséa, M.A., Brandão, C.R.F. 2013. Ant species (Hymenoptera, Formicidae) from the seasonally dry tropical forest of northeastern Brazil: a compilation from field surveys in Bahia and literature records. Revista Brasileira de Entomologia 57, 217–224 (doi:10.1590/s0085-56262013005000002).
- Wheeler, G. C.; Wheeler, J. 1952c. The ant larvae of the subfamily Ponerinae - Part II. Am. Midl. Nat. 48: 604-672 (page 651, larva described)
- Wheeler, W.M. 1928. Mermis parasitism and intercastes among ants. Journal of Experimental Zoology 50: 165-237 (doi:10.1002/jez.1400500202).
References based on Global Ant Biodiversity Informatics
- Andrade T., G. D. V. Marques, K. Del-Claro. 2007. Diversity of ground dwelling ants in Cerrado: an analysis of temporal variations and distinctive physiognomies of vegetation (Hymenoptera: Formicidae). Sociobiology 50(1): 121-134.
- Araujo Castilho G., F. Barbosa Noll, E. R. da Silva, and E. F. dos Santos. 2011. Diversidade de Formicidae (Hymenoptera) em um fragmento de Floresta Estacional Semidecídua no Noroeste do estado de São Paulo, Brasil. R. bras. Bioci., Porto Alegre 9(2): 224-230.
- Araujo L. M., and G. W. Fernandes. 2003. Altitudinal patterns in a tropical ant assemblage and variation in species richness between habitats. Lundiana 4(2): 103-109.
- Bezdeckova K., P. Bedecka, and I. Machar. 2015. A checklist of the ants (Hymenoptera: Formicidae) of Peru. Zootaxa 4020 (1): 101–133.
- Bieber A. G. D., O. P. G. Darrault, C. da Costa Ramos, K. K. Melo, and I. R. Leal. 2006. Formigas. p.244-262. In Porto K L, Tabarelli M, Almeida-Cortez J (eds) Diversidade biológica e conservação da Floresta Atlântica ao norte do rio São Francisco. Recife, Editora Universitária da UFPE, 363p
- Borgmeier T. 1923. Catalogo systematico e synonymico das formigas do Brasil. 1 parte. Subfam. Dorylinae, Cerapachyinae, Ponerinae, Dolichoderinae. Archivos do Museu Nacional (Rio de Janeiro) 24: 33-103.
- Brown W. L., Jr. 1976. Contributions toward a reclassification of the Formicidae. Part VI. Ponerinae, tribe Ponerini, subtribe Odontomachiti. Section A. Introduction, subtribal characters. Genus Odontomachus. Stud. Entomol. 19: 67-171.
- Bruch C. 1914. Catálogo sistemático de los formícidos argentinos. Revista del Museo de La Plata 19: 211-234.
- Caldart V. M., S. Iop, J. A. Lutinski, and F. R. Mello Garcia. 2012. Ants diversity (Hymenoptera, Formicidae) of the urban perimeter of Chapecó county, Santa Catarina, Brazil. Revista Brasileira de Zoociências 14 (1, 2, 3): 81-94.
- Calle Z., N. Henao-Gallego, C. Giraldo, and I. Armbrecht. 2013. A Comparison of Vegetation and Ground-dwelling Ants in Abandoned and Restored Gullies and Landslide Surfaces in the Western Colombian Andes. Restoration Ecology 21(6): 729-736.
- Cardoso, D.C., T.G. Sobrinho and J.H. Schoereder. 2010. Ant community composition and its relationship with phytophysiognomies in a Brazilian Restinga. Insectes Sociaux 57:293-301
- Chacon de Ulloa P., A. M. Osorio-Garica, R. Achury, and C. Bermudez-Rivas. 2012. Hormigas (Hymenoptera: Formicidae) del Bosque seco tropical (Bs-T) de la cuenca alta del rio Cauca, Colombia. Biota Colombiana 13(2): 165-181.
- Christianini A. V., A. J. Mayhé-Nunes, and P. S. Oliveira. 2012. Exploitation of Fallen Diaspores By Ants: Are There Ant-Plant Partner Choices? Biotropica 44: 360-367.
- Christianini A. V., and P. S. Oliveira. 2009. The relevance of ants as seed rescuers of a primarily bird-dispersed tree in the neotropical cerrado savanna. Oecologia 160: 735745.
- Christianini A. V., and P. S. Oliveira. 2013. Edge effects decrease ant-derived benefits to seedlings in a neotropical savanna. Arthropod-Plant Interactions 7: 191-199.
- Cividanes F. J., J. C. Barbosa, I. C. F. Martins, F. Pattaro, M. A. Nunes, R. Souza Santos. 2009. Diversity and spatial distribution of ground arthropods in agroecosystems. Bragantia, Campinas, 68(4): 991-1002.
- Clemes Cardoso D., and J. H. Schoereder. 2014. Biotic and abiotic factors shaping ant (Hymenoptera: Formicidae) assemblages in Brazilian coastal sand dunes: the case of restinga in Santa Catarina. Florida Entomologist 97(4): 1443-1450.
- Clemes Cardoso D., and M. Passos Cristiano. 2010. Myrmecofauna of the Southern Catarinense Restinga sandy coastal plain: new records of species occurrence for the state of Santa Catarina and Brazil. Sociobiology 55(1b): 229-239.
- Cuezzo, F. 1998. Formicidae. Chapter 42 in Morrone J.J., and S. Coscaron (dirs) Biodiversidad de artropodos argentinos: una perspectiva biotaxonomica Ediciones Sur, La Plata. Pages 452-462.
- Dattilo W. et al. 2019. MEXICO ANTS: incidence and abundance along the Nearctic-Neotropical interface. Ecology https://doi.org/10.1002/ecy.2944
- De la Mora, A., C. J. Murnen, and S. M. Philpott. 2013. Local and landscape drivers of ant-communities in Neotropical coffee landscapes. Biodiversity and Conservation 22: 871-888.
- De la Mora, A., G. Perez-Lachaud, J. P.Lachaud, and S. M. Philpott. 2015. Local and landscape drivers of ant parasitism in a coffee landscape. Environmental Entomology 44: 939-950.
- Delsinne T., Y. Roisin, and M. Leponce. 2007. Spatial and temporal foraging overlaps in a Chacoan ground-foraging ant assemblage. Journal of Arid Environments 71: 2944.
- Dias N. D. S., R. Zanetti, M. S. Santos, M. F. Gomes, V. Peñaflor, S. M. F. Broglio, and J. H. C. Delabie. 2012. The impact of coffee and pasture agriculture on predatory and omnivorous leaf-litter ants. Journal of Insect Science 13:29. Available online: http://www.insectscience.org/13.29
- Dias N. S., R. Zanetti, M. S. Santos, J. Louzada, and J. H. C. Delabie. 2008. Interaction between forest fragments and adjacent coffee and pasture agroecosystems: responses of the ant communities (Hymenoptera, Formicidae). Iheringia, Sér. Zool., Porto Alegre, 98(1): 136-142.
- Drose W., L. R. Podgaiski, C. Fagundes Dias, M. de Souza Mendonca. 2019. Local and regional drivers of ant communities in forest-grassland ecotones in South Brazil: A taxonomic and phylogenetic approach. Plos ONE 14(4): e0215310.
- Emery C. 1890. Voyage de M. E. Simon au Venezuela (Décembre 1887 - Avril 1888). Formicides. Annales de la Société Entomologique de France (6)10: 55-76.
- Emery C. 1896. Formiciden, gesammelt in Paraguay von Dr. J. Bohls. Zoologische Jahrbücher. Abteilung für Systematik, Geographie und Biologie der Tiere 9: 625-638.
- Emery C. 1906. Studi sulle formiche della fauna neotropica. XXVI. Bullettino della Società Entomologica Italiana 37: 107-194.
- Escalante Gutiérrez J. A. 1993. Especies de hormigas conocidas del Perú (Hymenoptera: Formicidae). Revista Peruana de Entomología 34:1-13.
- Escalante J. A. 1976. Hormigas del valle de K'Osnipata (Paucartambo, Cusco). Revista Peruana de Entomologia 107-108.
- Favretto M. A., E. Bortolon dos Santos, and C. J. Geuster. 2013. Entomofauna from West of Santa Catarina State, South of Brazil. EntomoBrasilis 6 (1): 42-63.
- Fernandes I., and J. de Souza. 2018. Dataset of long-term monitoring of ground-dwelling ants (Hymenoptera: Formicidae) in the influence areas of a hydroelectric power plant on the Madeira River in the Amazon Basin. Biodiversity Data Journal 6: e24375.
- Fernandes, P.R. XXXX. Los hormigas del suelo en Mexico: Diversidad, distribucion e importancia (Hymenoptera: Formicidae).
- Fernandez F. C., and L. Schneider S. 1989. Reconocimiento de hormigas en la reserva La Macarena. Revista Colombiana de Entomologia 15(1): 38-44.
- Fernández F. 2008. Subfamilia Ponerinae s.str. Pp. 123-218 in: Jiménez, E.; Fernández, F.; Arias, T.M.; Lozano-Zambrano F. H. (eds.) 2008. Sistemática, biogeografía y conservación de las hormigas cazadoras de Colombia. Bogotá: Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, xiv + 609 pp.
- Fernández F., and T. M. Arias-Penna. 2008. Las hormigas cazadoras en la región Neotropical. Pp. 3-39 in: Jiménez, E.; Fernández, F.; Arias, T.M.; Lozano-Zambrano, F. H. (eds.) 2008. Sistemática, biogeografía y conservación de las hormigas cazadoras de Colombia. Bogotá: Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, xiv + 609 pp.
- Fernández, F. and S. Sendoya. 2004. Lista de las hormigas neotropicales. Biota Colombiana Volume 5, Number 1.
- Fichaux M., B. Bechade, J. Donald, A. Weyna, J. H. C. Delabie, J. Murienne, C. Baraloto, and J. Orivel. 2019. Habitats shape taxonomic and functional composition of Neotropical ant assemblages. Oecologia 189(2): 501-513.
- Forel A. 1908. Catálogo systemático da collecção de formigas do Ceará. Boletim do Museu Rocha 1(1): 62-69.
- Forel A. 1909. Ameisen aus Guatemala usw., Paraguay und Argentinien (Hym.). Deutsche Entomologische Zeitschrift 1909: 239-269.
- Forel A. 1912. Formicides néotropiques. Part I. Annales de la Société Entomologique de Belgique. 56: 28-49.
- Franco W., N. Ladino, J. H. C. Delabie, A. Dejean, J. Orivel, M. Fichaux, S. Groc, M. Leponce, and R. M. Feitosa. 2019. First checklist of the ants (Hymenoptera: Formicidae) of French Guiana. Zootaxa 4674(5): 509-543.
- Gallardo A. 1918. Las hormigas de la República Argentina. Subfamilia Ponerinas. Anales del Museo Nacional de Historia Natural de Buenos Aires 30: 1-112.
- INBio Collection (via Gbif)
- Kempf W. W., and K. Lenko. 1976. Levantamento da formicifauna no litoral norte e ilhas adjacentes do Estado de São Paulo, Brasil. I. Subfamilias Dorylinae, Ponerinae e Pseudomyrmecinae (Hym., Formicidae). Studia Entomologica 19: 45-66.
- Kempf, W.W. 1972. Catalago abreviado das formigas da regiao Neotropical (Hym. Formicidae) Studia Entomologica 15(1-4).
- Kusnezov N. 1953. La fauna mirmecológica de Bolivia. Folia Universitaria. Cochabamba 6: 211-229.
- Kusnezov N. 1978. Hormigas argentinas: clave para su identificación. Miscelánea. Instituto Miguel Lillo 61:1-147 + 28 pl.
- Lapola D. M., and H. G. Fowler. 2008. Questioning the implementation of habitat corridors: a case study in interior São Paulo using ants as bioindicators. Braz. J. Biol., 68(1): 11-20.
- Leal I. R. and B. Cortes Lopes. 1992. Estrutura das comunidades de formigas (Hymenoptera: Formicidae) de solo e vegetacao no morro da lagoa de conceicao, Ilha de Santa Catarina, SC. Capa 5(1): 107-122.
- Levings S. C. 1983. Seasonal, annual, and among-site variation in the ground ant community of a deciduous tropical forest: some causes of patchy species distributions. Ecological Monographs 53(4): 435-455.
- Longino J. T. 2013. Ants of Nicargua. Consulted on 18 Jan 2013. https://sites.google.com/site/longinollama/reports/ants-of-nicaragua
- Longino J. T. L., and M. G. Branstetter. 2018. The truncated bell: an enigmatic but pervasive elevational diversity pattern in Middle American ants. Ecography 41: 1-12.
- Longino J. et al. ADMAC project. Accessed on March 24th 2017 at https://sites.google.com/site/admacsite/
- Lopes D. T., J. Lopes, I. Cardoso do Nascimento, and J. H. Delabie. 2010. Epigeic ants diversity (Hymenoptera, Formicidae) in three environments in Mata dos Godoy State Park, Londrina, State of Paraná, Brazil. Iheringia, Sér. Zool., Porto Alegre, 100(1): 84-90.
- Luederwaldt H. 1918. Notas myrmecologicas. Rev. Mus. Paul. 10: 29-64.
- Lutinski J. A., B. C. Lopes, and A. B. B.de Morais. 2013. Diversidade de formigas urbanas (Hymenoptera: Formicidae) de dez cidades do sul do Brasil. Biota Neotrop. 13(3): 332-342.
- Lutinski J. A., F. R. Mello Garcia, C. J. Lutinska, and S. Iop. 2008. Ants diversity in Floresta Nacional de Chapecó in Santa Catarina State, Brazil. Ciência Rural, Santa Maria 38(7): 1810-1816.
- Maes, J.-M. and W.P. MacKay. 1993. Catalogo de las hormigas (Hymenoptera: Formicidae) de Nicaragua. Revista Nicaraguense de Entomologia 23.
- Marinho C. G. S., R. Zanetti, J. H. C. Delabie, M. N. Schlindwein, and L. de S. Ramos. 2002. Ant (Hymenoptera: Formicidae) Diversity in Eucalyptus (Myrtaceae) Plantations and Cerrado Litter in Minas Gerais, Brazil. Neotropical Entomology 31(2): 187-195.
- Marques G. D. V., and K. Del-Claro. 2006. The Ant Fauna in a Cerrado area: The Influence of Vegetation Structure and Seasonality (Hymenoptera: Formicidae). Sociobiology 47(1): 1-18.
- Medeiros F. N. S., L. E. Lopes, P. R. S. Moutinho, P. S. Oliveira, and B. Hölldobler. 1992. Functional polygyny, agonistic interactions and reproductive dominance in the neotropical ant Odontomachus chelifer (Hymenoptera, Formicidae, Ponerinae). Ethology 91: 134-146.
- Medina U. C. A., F. Fernandez, and M. G. Andrade-C. 2010. Insectos: escarabajos coprofagos, hormigas y mariposas. Capitulo 6. Pp 197-215. En: Lasso, C. A., J. S. Usma, F. Trujillo y A. Rial (eds.). 2010. Biodiversidad de la cuenca del Orinoco: bases científicas para la identificación de áreas prioritarias para la conservación y uso sostenible de la biodiversidad. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, WWF Colombia, Fundación Omacha, Fundación La Salle e Instituto de Estudios de la Orinoquia (Universidad Nacional de Colombia). Bogotá, D. C., Colombia.
- Mentone T. O., E. A. Diniz, C. B. Munhae, O. C. Bueno, and M. S. C. Morini. 2011. Composition of ant fauna (Hymenoptera: Formicidae) at litter in areas of semi-deciduous forest and Eucalyptus spp., in Southeastern Brazil. Biota Neotrop. 11(2): http://www.biotaneotropica.org.br/v11n2/en/abstract?inventory+bn00511022011.
- Morini M. S. de C., C. de B. Munhae, R. Leung, D. F. Candiani, and J. C. Voltolini. 2007. Comunidades de formigas (Hymenoptera, Formicidae) em fragmentos de Mata Atlântica situados em áreas urbanizadas. Iheringia, Sér. Zool., Porto Alegre, 97(3): 246-252.
- Munhae C. B., Z. A. F. N. Bueno, M. S. C. Morini, and R. R. Silva. 2009. Composition of the Ant Fauna (Hymenoptera: Formicidae) in Public Squares in Southern Brazil. Sociobiology 53(2B): 455-472.
- Nascimento Santos M., J. H. C. Delabie, and J. M. Queiroz. 2019. Biodiversity conservation in urban parks: a study of ground-dwelling ants (Hymenoptera: Formicidae) in Rio de Janeiro City. Urban Ecosystems https://doi.org/10.1007/s11252-019-00872-8
- Oliveira Mentone T. de, E. A. Diniz, C. de Bortoli Munhae, O. Correa Bueno and M. S. de Castro Morini. 2012. Composition of ant fauna (Hymenoptera: Formicidae) at litter in areas of semi-deciduous forest and Eucalyptus spp., in Southeastern Brazil. Biota Neotrop 11(2): 237-246.
- Pacheco R., H. L. Vasconcelos, S. Groc, G. P. Camacho, and T. L. M. Frizzo. 2013. The importance of remnants of natural vegetation for maintaining ant diversity in Brazilian agricultural landscapes. Biodivers. Conserv. DOI 10.1007/s10531-013-0463-y
- Pacheco R., and H. L. Vasconcelos. 2007. Invertebrate conservation in urban areas: ants in the Brazilian Cerrado. Landscape and Urban Planning 81: 193199.
- Pacheco, R., R.R. Silva, M.S. de C. Morini, C.R.F. Brandao. 2009. A Comparison of the Leaf-Litter Ant Fauna in a Secondary Atlantic Forest with an Adjacent Pine Plantation in Southeastern Brazil. Neotropical Entomology 38(1):055-065
- Passos L., and P. S. Oliveira. 2002. Ants affect the distribution and performance of Clusia criuva seedlings, a primarily bird-dispersed rainforest tree. Journal of Ecology 90: 517-528.
- Passos, L. and P.S. Oliveira. 2002. Ants Affect the Distribution and Performance of Seedlings of Clusia criuva, a Primarily Bird-Dispersed Rain Forest Tree. Journal of Ecology 90(3):517-528.
- Passos, L. and P.S. Oliveira. 2003. Interactions between ants, fruits and seeds in a restinga forest in south-eastern Brazil. Journal of Tropical Ecology 19(3):261-270.
- Passos, L. and P.S. Oliviera. 2004. Interaction between Ants and Fruits of Guapira opposita (Nyctaginaceae) in a Brazilian Sandy Plain Rainforest: Ant Effects on Seeds and Seedling. Oecologia 139(3):376-382
- Pignalberi C. T. 1961. Contribución al conocimiento de los formícidos de la provincia de Santa Fé. Pp. 165-173 in: Comisión Investigación Científica; Consejo Nacional de Investigaciones Científicas y Técnicas (Argentina) 1961. Actas y trabajos del primer Congreso Sudamericano de Zoología (La Plata, 12-24 octubre 1959). Tomo III. Buenos Aires: Librart, 276 pp.
- Pires de Prado L., R. M. Feitosa, S. Pinzon Triana, J. A. Munoz Gutierrez, G. X. Rousseau, R. Alves Silva, G. M. Siqueira, C. L. Caldas dos Santos, F. Veras Silva, T. Sanches Ranzani da Silva, A. Casadei-Ferreira, R. Rosa da Silva, and J. Andrade-Silva. 2019. An overview of the ant fauna (Hymenoptera: Formicidae) of the state of Maranhao, Brazil. Pap. Avulsos Zool. 59: e20195938.
- Pizo M. A., P. R. Guimarães, Jr., and P. S. Oliveira. 2005. Seed removal by ants from faeces produced by different vertebrate species. Ecoscience 12: 136-140.
- Pizo M. A., and P. S. Oliveira. 2000. The Use of Fruits and Seeds by Ants in the Atlantic Forest of Southeast Brazil. Biotropica 32(b): 851-861.
- Pizo M. A., and P. S. Oliveira. 2001. Size and lipid content of nonmyrmecochorous diaspores: effects on the interaction with litter-foraging ants in the Atlantic rain forest of Brazil. Plant Ecology 157: 37-52.
- Pizo, M.A. and P.S. Oliviera. 1998. Interaction between ants and seeds of a nonmyrmecochorous neotropical tree, Cabralea canjerana (Meliaceae) in the Atlantic Forest of Southeast Brazil. American Journal of Botany 85(5):669-674.
- Pérez-Sánchez A. J., J. E. Lattke, and M. A. Riera-Valera. 2014. The Myrmecofauna (Hymenoptera: Formicidae) of the Macanao Semi-arid Peninsula in Venezuela: An Altitudinal Variation Glance. J Biodivers Biopros Dev 1: 116. doi:10.4172/ijbbd.1000116
- Raimundo R. L. G., A. V. L. Freitas, and P. S. Oliveira. 2009. Seasonal patterns in activity rhythm and foraging ecology in the Neotropical forest-dwelling ant, Odontomachus chelifer (Formicidae: Ponerinae). Annals of the Entomological Society of America 102: 1151-1157.
- Raimundo, R.L.G., A.V.L. Freitas and P.S. Oliviera.2009. Seasonal Patterns in Activity Rhythm and Foraging Ecology in the Neotropical Forest-Dwelling Ant, Odontomachus chelifer (Formicidae: Ponerinae). Ann. Entomol. Soc. Am. 102(6): 1151-1157
- Ramos L. S., R. Z. B. Filho, J. H. C. Delabie, S. Lacau, M. F. S. dos Santos, I. C. do Nascimento, and C. G. S. Marinho. 2003. Ant communities (Hymenoptera: Formicidae) of the leaf-litter in cerrado stricto sensu areas in Minas Gerais, Brazil. Lundiana 4(2): 95-102.
- Ramos L. de S., C. G. S. Marinho, R. Zanetti, and J. H. C. Delabie. 2003. Impacto de iscas formicidas granuladas sobre a mirmecofauna não-alvo em eucaliptais segundo duas formas de aplicacação / Impact of formicid granulated baits on non-target ants in eucalyptus plantations according to two forms of application. Neotropical Entomology 32(2): 231-237.
- Ramos L. de S., R. Zanetti, C. G. S. Marinho, J. H. C. Delabie, M. N. Schlindwein, and R. P. Almado. 2004. Impact of mechanical and chemical weedings of Eucalyptus grandis undergrowth on an ant community (Hymenoptera: Formicidae). Rev. Árvore 28(1): 139-146.
- Rodrigues de Souza D., E. Stingel, L. C. de Almeida, M. A. Lazarini, C. de Bortoli Munhae, A. J. Mayhe-Nunes, O. Correa Bueno, and M. Santina de C. Morini. 2010. Ant Diversity in a Sugarcane Culture without the Use of Straw Burning in Southeast, São Paulo, Brazil. American Journal of Agricultural and Biological Sciences 5 (2): 183-188.
- Rodrigues de Souza D., E. Stingel, L. C. de Almeida, M. A. Lazarini, C. de Bortoli Munhae, O. Correa Bueno, C. R. Archangelo, and M. Santina de C. Morini. 2010. Field methods for the study of ants in sugarcane plantations in Southeastern Brazil. Sci. Agric. (Piracicaba, Braz.) 67(6): 651-657.
- Rosa da Silva R. 1999. Formigas (Hymenoptera: Formicidae) do oeste de Santa Catarina: historico das coletas e lista atualizada das especies do Estado de Santa Catarina. Biotemas 12(2): 75-100.
- Rosa da Silva R., and B. Cortes Lopes. 1997. Ants (Hymenoptera: Formicidae) from Atlantic rainforest at Santa Catarina Island, Brazil: two years of sampling. Rev. Biol. Trop. 45(4): 1641-1648.
- Rosumek, F.B., M.A. Ulyssea, B.C. Lopes, J. Steiner. 2008. Formigas de solo e de bromélias em uma área de Mata Atlântica, Ilha de Santa Catarina, sul do Brasil: Levantamento de espécies e novos registros. Revista Biotemas 21(4):81-89.
- Salinas P. J. 2010. Catalogue of the ants of the Táchira State, Venezuela, with notes on their biodiversity, biogeography and ecology (Hymenoptera: Formicidae: Amblyioponinae, Ponerinae, Proceratiinae, Myrmicinae, Ecitoninae, Formicinae, Pseudomyrmecinae, Dolichoderinae). Boletín de la SEA 47: 315-328.
- Santoandre S., J. Filloy, G. A. Zurita, and M. I. Bellocq. 2019. Ant taxonomic and functional diversity show differential response to plantation age in two contrasting biomes. Forest Ecology and Management 437: 304-313.
- Santos M. S., J. N. C. Louzada, N. Dias, R. Zanetti, J. H. C. Delabie, and I. C. Nascimento. 2006. Litter ants richness (Hymenoptera, Formicidae) in remnants of a semi-deciduous forest in the Atlantic rain forest, Alto do Rio Grande region, Minas Gerais, Brazil. Iheringia, Sér. Zool., Porto Alegre, 96(1): 95-101.
- Santos-Junior L. C., J. M. Saraiva, R. Silvestre, and W. F. Antonialli-Junior. 2014. Evaluation of Insects that Exploit Temporary Protein Resources Emphasizing the Action of Ants (Hymenoptera, Formicidae) in a Neotropical Semi-deciduous Forest. Sociobiology 61(1): 43-51
- Santschi F. 1922. Description de nouvelles fourmis de l'Argentine et pays limitrophes. Anales de la Sociedad Cientifica Argentina. 94: 241-262.
- Schmidt K., R. Corbetta, and A.J. Aires de Carmargo. 2005. Formigas (Hymenoptera: Formicidae) da Ilha João da Cunha, SC: composição e diversidade. Biotemas, 18 (1): 57 - 71
- Schütte M. S., J. M. Queiroz, A. J. Mayhé-Nunes, M. P. dos S. Pereira. 2007. Structured inventory of ants (Hymenoptera, Formicidae) in atlantic slope rain-forest of Marambaia Island, RJ. Iheringia, Sér. Zool., Porto Alegre 97(1): 103-110.
- Silva R. R., R. S. Machado Feitosa, and F. Eberhardt. 2007. Reduced ant diversity along a habitat regeneration gradient in the southern Brazilian Atlantic Forest. Forest Ecology and Management 240: 61-69.
- Silva R.R., and C. R. F. Brandao. 2014. Ecosystem-Wide Morphological Structure of Leaf-Litter Ant Communities along a Tropical Latitudinal Gradient. PLoSONE 9(3): e93049. doi:10.1371/journal.pone.0093049
- Silvestre R., C. R. F. Brandão, and R. R. Silva da 2003. Grupos funcionales de hormigas: el caso de los gremios del cerrado. Pp. 113-148 in: Fernández, F. (ed.) 2003. Introducción a las hormigas de la región Neotropical. Bogotá: Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, xxvi + 424 pp.
- Silvestre R., M. F. Demetrio, and J. H. C. Delabie. 2012. Community Structure of Leaf-Litter Ants in a Neotropical Dry Forest: A Biogeographic Approach to Explain Betadiversity. Psyche doi:10.1155/2012/306925
- Sobrinho T. G., and J. H. Schoereder. 2007. Edge and shape effects on ant (Hymenoptera: Formicidae) species richness and composition in forest fragments. Biodivers Conserv 16: 14591470.
- Sobrinho T., J. H. Schoereder, C. F. Sperber, and M. S. Madureira. 2003. Does fragmentation alter species composition in ant communities (Hymenoptera: Formicidae)? Sociobiology 42(2): 329-342.
- Suguituru S. S., M. Santina de Castro Morini, R. M. Feitosa, and R. Rosa da Silva. 2015. Formigas do Alto Tiete. Canal 6 Editora 458 pages
- Ulloa Chacon P., M. L. Baena, J. Bustos, R. C. Aldana, J. A. Aldana, and M. A. Gamboa. 1996. Fauna de hormigas del departamento del Valle del Cauca (Colombia). Pp. 413-451. In Andrade-C M. G., G. Amat Garcia, and F. Fernandez. Insectoss de Colombia, estudios escogidos.
- Ulyssea M.A., C. E. Cereto, F. B. Rosumek, R. R. Silva, and B. C. Lopes. 2011. Updated list of ant species (Hymenoptera, Formicidae) recorded in Santa Catarina State, southern Brazil, with a discussion of research advances and priorities. Revista Brasileira de Entomologia 55(4): 603-611.
- Vittar, F. 2008. Hormigas (Hymenoptera: Formicidae) de la Mesopotamia Argentina. INSUGEO Miscelania 17(2):447-466
- Vittar, F., and F. Cuezzo. "Hormigas (Hymenoptera: Formicidae) de la provincia de Santa Fe, Argentina." Revista de la Sociedad Entomológica Argentina (versión On-line ISSN 1851-7471) 67, no. 1-2 (2008).
- Vásquez-Bolaños M. 2011. Lista de especies de hormigas (Hymenoptera: Formicidae) para México. Dugesiana 18: 95-133
- Wheeler W. M. 1916. Ants collected in British Guiana by the expedition of the American Museum of Natural History during 1911. Bulletin of the American Museum of Natural History 35: 1-14.
- Wheeler W. M. 1925. Neotropical ants in the collections of the Royal Museum of Stockholm. Arkiv för Zoologi 17A(8): 1-55.
- Wild, A. L. "A catalogue of the ants of Paraguay (Hymenoptera: Formicidae)." Zootaxa 1622 (2007): 1-55.
- da Silva, R.R., C.R.F. Brandao, and R. Silvestre. 2004. Similarity Between Cerrado Localities in Central and Southeastern Brazil Based on the Dry Season Bait Visitors Ant Fauna. Studies on Neotropical Fauna and Environment 39(3):191-199.
- de Miranda M., V. Barbosa de Andrade, G. D. Vieira Marques, and V. S. Sul Moreira. 2006. Ant fauna (Formicidae, Hymenoptera) in an urban fragment area of semideciduous mesophitic forest. Zoociencias 8(1): 49-54.
- de Souza D. R., S. G. dos Santos, C. de B. Munhae, and M. S. de C. Morini. 2012. Diversity of Epigeal Ants (Hymenoptera: Formicidae) in Urban Areas of Alto Tietê. Sociobiology 59(3): 703-117.
- Pages using DynamicPageList3 parser function
- Polygynous
- North subtropical
- Tropical
- South subtropical
- South temperate
- Eucharitid wasp Associate
- Host of Latina rugosa
- Phorid fly Associate
- Host of Apocephalus catholicus
- Host of Apocephalus paldiae
- Host of Dohrniphora sp. (not D. longi-gp)
- Host of Dohrniphora sp.
- Nematode Associate
- Host of Mermithidae (unspecified "Mermix")
- Karyotype
- Species
- Extant species
- Formicidae
- Ponerinae
- Ponerini
- Odontomachus
- Odontomachus chelifer
- Ponerinae species
- Ponerini species
- Odontomachus species
- Need Body Text