Tetramorium caespitum
| Tetramorium caespitum | |
|---|---|

| |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Arthropoda |
| Class: | Insecta |
| Order: | Hymenoptera |
| Family: | Formicidae |
| Subfamily: | Myrmicinae |
| Tribe: | Crematogastrini |
| Genus: | Tetramorium |
| Species: | T. caespitum |
| Binomial name | |
| Tetramorium caespitum (Linnaeus, 1758)
| |
| Subspecies | |
| |
| Synonyms | |
| |


This is the most common European Tetramorium species. Long believed to be introduced to North America, Wagner et al. (2017) conclusively determined the North American species is Tetramorium immigrans.
Wagner et al. (2017) - Moderately thermophilic, TAS of 465 sites 16.1 ± 2.0 °C [7.9, 21.1], different from all species except Tetramorium indocile. Most common species in most of Europe and more euryoecious than other species of complex. Most records from non-forested habitats like meadows, pastures, heaths, arid or semi-arid grasslands, vineyards, fallow grounds, ruderal areas, road embankments, rock heaps, gravel pits, river banks, but also light pine and oak forests. Urban areas like parks, pavements, and roadsides. Nest construction more flexible than in other species: in soil, under stones, rarely in dead wood; only species building soil mounds higher than 10 cm.
Pashaei Rad et al. (2018) found this species in Iran in Caspian moist littoral and moderate rainfall montane areas.
A recent revision of the Tetramorium caespitum complex (Wagner et al. 2017) showed that the group contains several cryptic species, thus all old Greek records of Tetramorium caespitum need confirmation. Borowiec & SaLata (2019c) confirmed its occurrence in Epirus, Macedonia, Peloponnese and Thessaly, and in this paper its presence is confirmed from Sterea Ellas. In Peloponnese and Aetolia-Acarnania, its nests were observed in mountain pastures and fir forest (Borowiec & Salata, 2021).
Photo Gallery
 Tetramorium atratulum dealate gyne moving freely in Tetramorium caespitum nest associated with the social parasite Strongylognathus testaceus (photo: A. Purkart, from Purkart et al., 2021, Fig. 8).
Tetramorium atratulum dealate gyne moving freely in Tetramorium caespitum nest associated with the social parasite Strongylognathus testaceus (photo: A. Purkart, from Purkart et al., 2021, Fig. 8).


_Fig_8.jpg)
{kind=link}
Identification
A member of the Tetramorium caespitum species complex. See Wagner et al. (2017) and https://webapp.uibk.ac.at/ecology/tetramorium/ for keys.
Siberian Species
Tetramorium sibiricum differs from Tetramorium indocile in having a longer scape, a more developed sculpture on postocular head sides, a narrower petiole and postpetiole, longer pronotal setae and a longer mesosoma and eye. The main differences to Tetramorium caespitum are the narrower petiole and postpetiole, the more developed sculpture and microsculpture on postocular head sides and on dorsum of 1st gaster tergite, the higher postpetiole, the longer scape and eye and a longer distance from spine tip to dorsocaudal corner of metapleural lobe.
Three species of the T. caespitum complex occur in Siberia west and north of the Reinig Line. This faunal divide separates East Siberian, Inner Mongolian, Chinese and Tibetan species from those of Central Siberia, West Siberia and the Turanian region (DE LATTIN, 1967).
The methodology of species identification applied by Seifert (2021) is extremely complex and data recording is by far the most time-consuming of any ant genus investigated so far by the author – a challenge even for experienced and specialized investigators having access to adequate equipment. An attempt to simplify the separation of the three West and Central Siberian species was done in writing a key that uses absolute primary data (i. e. without RAV-correction) and a strongly reduced character set.
1. Postoculo-temporal area of head with rather many costae and costulae, POTCos 11.4 ± 2.1. Anterior dorsum of 1st gaster tergite with more complex elements of stickman-like or reticulate microsculpture, MC1TG 16.6 ± 3.2. Propodeal spines and eye slightly longer. With all linear measurements given in mm, discriminant function 0.183*POTCos + 0.12*MC1TG – 34.79*FL + 46.67*EL + 17.54*ML + 28.28*MPSP – 99.07*PeW – 3.793 > 0 [error 0 % in 21 specimens] => Tetramorium sibiricum
– Postoculo-temporal area of head with fewer costae and costulae, POTCos 8.4 ± 2.4. Anterior dorsum of 1st gaster tergite with less complex elements of stickman-like or reticulate microsculpture, MC1TG 11.2 ± 3.0. Propodeal spines and eye slightly shorter. Discriminant < 0 [error 0 % in 338 specimens] => 2
2. Longest hair near anterolateral pronotal corner shorter, PnHL 0.168 ± 0.020 mm. Anterior dorsum of 1st gaster tergite with more complex elements of stickman-like microsculpture, MC1TG 13.5 ± 2.8. With all linear measurements given in mm, discriminant 0.127*POTCos – 0.173*MC1TG + 85.54*SWd – 49.08*MPSP + 62.52*MPPL + 21.985*PnHL – 9.026 < 0 [error 9.7% in 72 specimens] => Tetramorium indocile
– Longest hair near anterolateral pronotal corner longer, PnHL 0.212 ± 0.022 mm. Anterior dorsum of 1st gaster tergite with only scattered and less complex elements of stickman-like microsculpture, MC1TG 10.6 ± 2.7. Discriminant > 0 [error 1.9 % in 266 specimens] => Tetramorium caespitum
Keys including this Species
- Key to US Tetramorium species
- Key to Tetramorium of India
- Key to Afrotropical Tetramorium species groups
Distribution
Europe, up to 63° N, Caucasus. This species does not occur in North America (Wagner et al. 2017).
Latitudinal Distribution Pattern
Latitudinal Range: 65.505015° to 27.766667°.
| North Temperate |
North Subtropical |
Tropical | South Subtropical |
South Temperate |
- Source: AntMaps
Distribution based on Regional Taxon Lists
Palaearctic Region: Afghanistan, Albania, Armenia, Austria, Azerbaijan, Balearic Islands, Belarus, Belgium, Bulgaria, Canary Islands, Channel Islands, China, Croatia, Democratic Peoples Republic of Korea, Denmark, Estonia, Finland, France, Georgia, Germany, Gibraltar, Hungary, Iberian Peninsula, Iran, Israel, Italy, Jersey, Kazakhstan, Kyrgyzstan, Latvia, Liechtenstein, Lithuania, Luxembourg, Malta, Monaco, Montenegro, Netherlands, Norway, Poland, North Macedonia, Republic of Moldova, Romania, Russian Federation, Slovakia, Spain, Sweden, Switzerland, Türkiye, Turkmenistan, Ukraine, United Kingdom of Great Britain and Northern Ireland.
Distribution based on AntMaps
Distribution based on AntWeb specimens
Check data from AntWeb
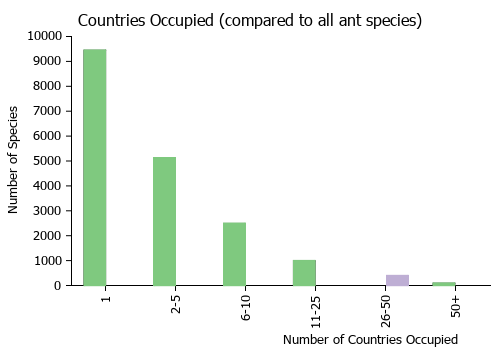
Countries Occupied
| Number of countries occupied by this species based on AntWiki Regional Taxon Lists. In general, fewer countries occupied indicates a narrower range, while more countries indicates a more widespread species. |
|
Biology
Monogynous (Seifert 2007). Hybridizes with Tetramorium immigrans (Wagner et al. 2017; Cordonnier et al. 2019).
Adult sexuals in nests on 25 June ± 14d [28 May, 19 August] (n = 67). Direct swarming behavior observed on 14 June, 15 June at 11:05 true solar time, and 30 June at 07:15 true solar time.
Collingwood (1979) - The species tends to be coastal in North Europe but also inland on heath and on the open borders of woodland, nesting in the earth and also under stones. Colonies are normally single queened, but populous with up to 10,000 or more workers. This species is moderately aggressive, living by predation on other arthropods, scavenging and also from root aphid honeydew. Seeds of various herbs and grasses are often collected into the nest. The alatae are conspicuously large compared with the workers; they are developed in the early summer and fly in late June and July.
Hybridizes with Tetramorium immigrans (Wagner et al. 2017; Cordonnier et al. 2019; Cordonnier et al. 2020).
Flight Period
| X | X | X | |||||||||
| Jan | Feb | Mar | Apr | May | Jun | Jul | Aug | Sep | Oct | Nov | Dec |
Source: antkeeping.info.
- Check details at Worldwide Ant Nuptial Flights Data, AntNupTracker and AntKeeping.
 Explore: Show all Flight Month data or Search these data. See also a list of all data tables or learn how data is managed.
Explore: Show all Flight Month data or Search these data. See also a list of all data tables or learn how data is managed.
Association with Other Organisms
- Explore: Show all Associate data or Search these data. See also a list of all data tables or learn how data is managed.
Other Ants
- This ant is parasitized by the workerless inquilines Strongylognathus karawajewi (de la Mora et al., 2021; Sanetra & Buschinger, 2000), Tetramorium atratulum (Seifert, 2018; Wagner et al., 2017) and Tetramorium inquilinum, the inquiline Strongylognathus testaceus (de la Mora et al., 2021; Novák & Sadil, 1941; Sanetra et al., 1994; Sanetra et al., 1999; Seifert, 1996; Sanetra & Buschinger, 2000; Seifert, 2018), and the slave-makers Strongylognathus christophi , Strongylognathus huberi (de la Mora et al., 2021; Sanetra & Buschinger, 2000; Seifert, 2018), Strongylognathus italicus (de la Mora et al., 2021; Seifert, 2018) and Strongylognathus kratochvili (de la Mora et al., 2021; Seifert, 2018).
- This species is a xenobiont for the ant Formica fusca (a xenobiont) in Poland (Kanizsai et al., 2013; Włodarczyk et al., 2009) (Clearcut of managed forest, in tree stumps).
- This species is a xenobiont for the ant Formicoxenus nitidulus (a xenobiont) (Martin et al. 2007; Wilson 1971).
- This species is a xenobiont for the ant Lasius flavus (a xenobiont) in Finland (Czechowski, 2004; Kanizsai et al., 2013) (Different successional series of rocky habitats. Under stone).
Hemiptera
- This species is associated with the aphids Aphis fabae, Aphis farinosa, Aphis gossypii, Aphis salviae, Aphis spiraecola, Brachycaudus cardui, Macrosiphoniella sanborni and Myzus lythri (Saddiqui et al., 2019 and included references)
Nematodes
- This species is a host for the nematode Mermithidae (unspecified "Mermix") (a parasite) in Germany (Wuerzburg) (Gösswald, 1938; Laciny, 2021).
- This species is a host for the nematode Mermithidae (unspecified "Mermix") (a parasite) in Switzerland (Wallis) (Kutter, 1958; Laciny, 2021).
- This species is a host for the nematode Oscheius dolichura (a parasite) (Wahab, 1962).
- This species is a host for the nematode Oscheius dolichura (a parasite) (Quevillon, 2018) (multiple encounter modes; indirect transmission; transmission outside nest).
Mites
- This species is a host for the mite Petalomium nataliae (a parasite) in Iran (Rahiminejad & Hajiqanbar, 2020) (ectoparasite).
- This species is a host for the mite Scutacarus longisetus (a parasite) (Khaustov, 2015) (ectoparasite).
Cestoda
- This species is a host for the cestode Raillietina caucasica (a parasitoid) (Quevillon, 2018) (encounter mode secondary; indirect transmission; transmission outside nest).
- This species is a host for the cestode Raillietina circumvalata (a parasitoid) (Quevillon, 2018) (encounter mode secondary; indirect transmission; transmission outside nest).
- This species is a host for the cestode Raillietina echinobothrida (a parasitoid) (Quevillon, 2018) (encounter mode secondary; indirect transmission; transmission outside nest).
- This species is a host for the cestode Raillietina fedjuschini (a parasitoid) (Quevillon, 2018) (encounter mode secondary; indirect transmission; transmission outside nest).
- This species is a host for the cestode Raillietina tetragona (a parasitoid) (Quevillon, 2018) (encounter mode secondary; indirect transmission; transmission outside nest).
Fungi
- This species is a host for the fungus Myrmicinosporidium durum (a parasitoid) (Quevillon, 2018) (encounter mode primary; direct transmission; transmission within nest).
- This species is a host for the fungus Tarichium sp. (a parasitoid) (Quevillon, 2018) (encounter mode primary; direct transmission; transmission outside nest).
- This species is a host for the fungus Myrmicinosporidium durum (a pathogen) (Espadaler & Santamaria, 2012).
Life History Traits
- Queen number: monogynous (Frumhoff & Ward, 1992)
- Mean colony size: 14,000 (Brian et al., 1967; Pasteels et al., 1987; Beckers et al., 1989)
- Foraging behaviour: mass recruiter (Brian et al., 1967; Pasteels et al., 1987; Beckers et al., 1989)
Castes
Worker
Images from AntWeb
   
| |
| Worker. Specimen code casent0173203. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by CAS, San Francisco, CA, USA. |
Queen

| |
| . | |
Male
Images from AntWeb
    
| |
| Male (alate). Specimen code casent0178769. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by CAS, San Francisco, CA, USA. |
Nomenclature
The following information is derived from Barry Bolton's Online Catalogue of the Ants of the World.
- caespitum. Formica caespitum Linnaeus, 1758: 581 (w.) EUROPE. Latreille, 1798: 50 (q.m.); Mayr, 1861: 62 (q.m.); Wheeler, G.C. & Wheeler, J. 1954d: 445 (l.); Hauschteck, 1961: 221 (k.); Imai, 1966: 119 (k.). Combination Manica: Jurine, 1807: 279; in Tetramorium: Mayr, 1855: 426. Senior synonym of fuscula: Smith, F. 1851: 118, Radchenko, 2007: 31; of modesta Foerster: Curtis, 1854: 215; Mayr, 1855: 426; of fusca: Dalla Torre, 1893: 132; of transversinodis: Brown, 1949a: 47; of immigrans: Bolton, 1979: 171; of himalayanum, indocile, transbaicalense: Radchenko, 1992b: 50; of hammi: Bolton, 1995b: 405; of jiangxiense: Wu & Wang, 1995: 82; of fusciclavum: Sanetra, Güsten & Schulz, 1999: 320. Current subspecies: nominal plus barabense, caespitomoravicum, flavidulum, japonicum, pallidum, typicum. See also: Emery, 1909d: 697; Bondroit, 1918: 107; Emery, 1925c: 177; Baroni Urbani, 1971c: 135; Kutter, 1977c: 157; Arnol'di & Dlussky, 1978: 544; Smith, D.R. 1979: 1400; Collingwood, 1979: 84; Cammaerts, Pasteels et al. 1985: 109; Kupyanskaya, 1990: 151; López, 1991a: 31; López, 1991b: 73; López, et al. 1992: 169; Radchenko, Czechowski & Czechowska, 1998: 108.
- fusca. Formica fusca Leach, 1825: 290 (q.m.) FRANCE. [Unresolved junior primary homonym of Formica fusca Linnaeus, 1758: 580.] Junior synonym of caespitum: Dalla Torre, 1893: 132.
- fuscula. Myrmica fuscula Nylander, 1846a: 935, pl. 18, figs. 34, 36 (w.q.m.) FINLAND. Junior synonym of caespitum: Smith, F. 1851: 118; Radchenko, 2007: 31.
- modesta. Myrmica modesta Foerster, 1850a: 49 (w.) GERMANY. Junior synonym of caespitum: Curtis, 1854: 215; Mayr, 1855: 426.
- himalayanum. Tetramorium caespitum subsp. himalayanum Viehmeyer, 1914b: 38 (w.q.m.) INDIA. Junior synonym of caespitum: Radchenko, 1992b: 50.
- hammi. Tetramorium caespitum var. hammi Donisthorpe, 1915d: 178 (w.) GREAT BRITAIN. Junior synonym of caespitum: Bolton, 1995b: 408.
- immigrans. Tetramorium caespitum var. immigrans Santschi, 1927a: 54 (w.) CHILE. Junior synonym of caespitum: Bolton, 1979: 171.
- transbaicalense. Tetramorium semilaeve subsp. transbaicalense Ruzsky, 1936: 93 (w.) KAZAKHSTAN. Junior synonym of caespitum: Radchenko, 1992b: 50.
- transversinodis. Myrmica (Myrmica) brevinodis var. transversinodis Enzmann, J. 1946b: 47, figs. 1,2 (w.) U.S.A. Junior synonym of caespitum: Brown, 1949a: 47.
- fusciclavum. Tetramorium caespitum var. fusciclavum Consani & Zangheri, 1952: 42 (w.) ITALY. [First available use of Tetramorium caespitum subsp. caespitum var. fusciclava Emery, 1925c: 187; unavailable name (Bolton, 1995b: 408).] Junior synonym of caespitum: Sanetra, Güsten & Schulz, 1999: 320.
- jiangxiense. Tetramorium jiangxiense Wang & Xiao, in Wang, M., Xiao & Wu, 1988: 269, figs. 24, 25 (w.) CHINA. Junior synonym of caespitum: Wu & Wang, 1995: 82.
Type Material
Wagner et al. (2017) - Neotype designation: Schlick-Steiner et al. 2006. Floghult Bohuslan (Sweden), 58.97° N, 11.42° E, 100 m a.s.l., leg. C.A. Collingwood, 21.VI. 2000. Unless otherwise noted the text for the remainder of this section is reported from the publication that includes the original description.
Description
Worker
Wagner et al. (2017) - Larger than most species of complex, CS = 761 ± 50 [591, 867] μm. Dark brown to blackish.
Head moderately elongate, CL / CW = 1.012 ± 0.015 [0.969, 1.043]. Eye rather small, EYE / CS = 0.171 ± 0.005 [0.158, 0.188]. Scape length moderate, SLd / CS = 0.777 ± 0.015 [0.724, 0.812]. Mesosoma long and wide, ML / CS = 1.172 ± 0.026 [1.104, 1.233], MW / CS = 0.645 ± 0.015 [0.605, 0.687].
Promesonotal dorsum convex, metanotal groove shallow. – Head dorsum and occiput with longitudinal costae and costulae, in Iberia longitudinal costae and costulae of head dorsum sometimes interrupted by smooth and shiny areas. Postoculo-temporal area of head with moderate number of longitudinal costae and costulae, POTCos = 7.45 ± 1.92 [3.38, 12.13]. Mesosoma dorsum longitudinally rugulose, in Iberia longitudinal costae and costulae sometimes interrupted by smooth and shiny areas. Lateral side of propodeum with a moderately pronounced smooth and shiny area, Ppss = 39.9 ± 20.0 [13.3, 107.7]. Dorsum of petiolar node smooth or with slightly microreticulate sculpture. General surface appearance on average moderately smooth and shiny compared with other species. – Connected stickman- like or reticulate microsculpture: small units scattered over 1st gastral tergite, MC1TG = 12.62 ± 2.31 [7.00, 19.58]. – Some workers with long c-shaped, crinkly, or sinuous hairs on ventral head posterior to buccal cavity.
Male
Wagner et al. (2017) - Paramere structure belongs to caespitum-like form: ventral paramere lobe with one or two sharp corners; without distinct emargination between paramere lobes in posterior view, both paramere lobes reduced in size; in ventro-posterior view, second corner on ventral paramere lobe missing or < 87 μm apart from first. In posterior view, typically only one sharp corner on ventral lobe.
Karyotype
- See additional details at the Ant Chromosome Database.
- Explore: Show all Karyotype data or Search these data. See also a list of all data tables or learn how data is managed.
- n = 14, 2n = 28 (Japan) (Imai, 1966; Imai, 1969; Imai & Yosida, 1964).
- n = 14, karyotype = 5SM+2ST+7A (Spain) (Palomeque et al., 1987; Palomeque et al., 1990b; Lorite et al., 2000).
- n = 14, 2n = 28 (Switzerland) (Hauschteck, 1961).
Worker Morphology
- Explore: Show all Worker Morphology data or Search these data. See also a list of all data tables or learn how data is managed.
- Caste: monomorphic
References
- Arcos, J., Chaves, D., Alarcón, P., Rosado, A. 2022. First record of Temnothorax convexus (Forel, 1894) in Portugal (Hymenoptera: Formicidae) with an updated checklist of the ants from the country. Sociobiology, 69(2), e7623 (doi:10.13102/sociobiology.v69i2.7623).
- Arcos, J., Chaves, D., Alarcón, P., Rosado, Á. 2022. First record of Temnothorax convexus (Forel, 1894) in Portugal (Hymenoptera: Formicidae) with an updated checklist of the ants from the country. Sociobiology, 692), e7623 (doi:10.13102/sociobiology.v69i2.7623).
- Arnol'di, K. V.; Dlussky, G. M. 1978. Superfam. Formicoidea. 1. Fam. Formicidae - ants. Pp. 519-556 in: Medvedev, G. S. (ed.) Keys to the insects of the European part of the USSR. Vol. 3. Hymenoptera. Part 1. Opredeliteli Faune SSSR 119:3-584. (page 544, see also)
- Baroni Urbani, C. 1971c. Catalogo delle specie di Formicidae d'Italia (Studi sulla mirmecofauna d'Italia X). Memorie della Societa Entomologica Italiana 50: 5-287 (page 135, see also)
- Beckers R., Goss, S., Deneubourg, J.L., Pasteels, J.M. 1989. Colony size, communication and ant foraging Strategy. Psyche 96: 239-256 (doi:10.1155/1989/94279).
- Bharti, H. & Kumar, R. 2012. Taxonomic studies on genus Tetramorium Mayr (Hymenoptera, Formicidae) with report of two new species and three new records including a tramp species from India with a revised key. ZooKeys. 207:11-35. doi:10.3897/zookeys.207.3040
- Bolton, B. 1979. The ant tribe Tetramoriini (Hymenoptera: Formicidae). The genus Tetramorium Mayr in the Malagasy region and in the New World. Bull. Br. Mus. (Nat. Hist.) Entomol. 38: 129-181 (page 171, Senior synonym of immigans)
- Bolton, B. 1995b. A new general catalogue of the ants of the world. Cambridge, Mass.: Harvard University Press, 504 pp. (page 405, Senior synonym of hammi; new synonymy)
- Bondroit, J. 1918. Les fourmis de France et de Belgique. Ann. Soc. Entomol. Fr. 87: 1-174 (page 107, see also)
- Boomsma, J.J., Leusink, A. 1981. Weather conditions during nuptial flights of four European ant species. Oecologia 50, 236–241. (doi:10.1007/bf00348045).
- Borowiec, L. 2014. Catalogue of ants of Europe, the Mediterranean Basin and adjacent regions (Hymenoptera: Formicidae). Genus (Wroclaw) 25(1-2): 1-340.
- Borowiec, L., Salata, S. 2018. Tetramorium immigrans Santschi, 1927 (Hymenoptera: Formicidae) nowy gatunek potencjalnie inwazyjnej mrówki w polsce. Acta entomologica silesiana 26:1-5 (doi:10.5281/ZENODO.1169156).
- Borowiec, L., Salata, S. 2020. Review of ants (Hymenoptera: Formicidae) from Jordan. Annals of the Upper Silesian Museum in Bytom, Entomology 29 (online 2): 1-26 (doi:10.5281/zenodo.3733156).
- Borowiec, L., Salata, S. 2021. Notes on ants (Hymenoptera: Formicidae) from Western Greece. Annals of the Upper Silesian Museum in Bytom Entomology 30: 1-23 (doi:10.5281/ZENODO.5571258).
- Borowiec, L., Salata, S. 2022. Notes on ants (Hymenoptera: Formicidae) of Thassos Island, Greece. Annals of the Upper Silesian Museum in Bytom, Entomology 31 (online 2): 1-15 (doi:10.5281/ZENODO.6123287).
- Borowiec, L., van Delft, J.P.L., van Delft, J.J.C.W., Salata, S. 2023. Five ant species (Hymenoptera: Formicidae) new to the Greek fauna with notes on ants from Greek Thrace. Annales of the Upper Silesian Museum in Bytom, Entomology 32 (online 008), 1-13 (doi:10.5281/ZENODO.10101028).
- Bračko, G. 2019. New data on the ant fauna (Hymenoptera: Formicidae) of Azerbaijan. Caucasian Entomological Bulletin 15, 165–175 (doi:10.23885/181433262019151-165175).
- Bracko, G., Wagner, H.C., Schulz, A., Gioahin, E., Maticic, J., Trantnik, A. 2014. New investigation and a revised checklist of the ants (Hymenoptera: Formicidae) of the Republic of Macedonia. North-Western Journal of Zoology 10: 10-24.
- Brown, W. L., Jr. 1949a. Synonymic and other notes on Formicidae (Hymenoptera). Psyche (Camb.) 56: 41-49 (page 47, Senior synonym of transversinodis)
- Bulter, I. 2020. Hybridization in ants. Ph.D. thesis, Rockefeller University.
- Buschinger, A. 1987. Teleutomyrmex schneideri Kutter 1950 and other parasitic ants found in the Pyrenees (Hymenoptera, Formicidae). Spixiana 10, 81-83.
- Carroll, T.M. 2011. The ants of Indiana (Hymenoptera: Formicidae). M.S. thesis, Purdue University.
- Castracani, C., Spotti, F.A., Schifani, E., Giannetti, D., Ghizzoni, M., Grasso, D.A., Mori, A. 2020. Public engagement provides first insights on Po Plain ant communities and reveals the ubiquity of the cryptic species Tetramorium immigrans (Hymenoptera, Formicidae). Insects 11, 678. (doi:10.3390/insects11100678).
- Chalissery, J.M., Gries, R., Alamsetti, S.K., Ardiel, M.J., Gries, G. 2021. Identification of the trail pheromone of the Pavement Ant Tetramorium immigrans (Hymenoptera: Formicidae). Journal of Chemical Ecology (doi:10.1007/s10886-021-01317-3).
- Collingwood, C. A. 1979. The Formicidae (Hymenoptera) of Fennoscandia and Denmark. Fauna Entomol. Scand. 8: 1-174 (page 84, see also)
- Cordonnier, M., Escarguel, G., Dumet, A., Kaufmann, B. 2020. Multiple mating in the context of interspecific hybridization between two Tetramorium ant species. Heredity 124, 675–684 (doi:10.1038/s41437-020-0310-3).
- Cordonnier, M., Gayet, T., Escarguel, G., Kaufmann, B. 2019. From hybridization to introgression between two closely related sympatric ant species. Journal of Zoological Systematics and Evololutionary Research 2019;00:1–11 (DOI 10.1111/jzs.12297).
- Csata, E., Dussutour, A. 2019. Nutrient regulation in ants (Hymenoptera: Formicidae): a review. Myrmecological News 29: 111-124 (doi:10.25849/MYRMECOL.NEWS_029:111).
- Csosz, S. ; Marko, B. 2004. Redescription of Tetramorium hungaricum Roeszler, 1935, a related species of T. caespitum (Linnaeus, 1758) (Hymenoptera: Formicidae). Myrmecologische Nachrichten 6: 49-59 (page 56, diagnostic characters)
- Csősz, S., Báthori, F., Gallé, L., Lőrinczi, G., Maák, I., Tartally, A., Kovács, É., Somogyi, A.Á., Markó, B. 2021. The myrmecofauna (Hymenoptera: Formicidae) of Hungary: Survey of ant species with an annotated synonymic inventory. Insects 16;12(1):78 (doi:10.3390/insects12010078).
- Csosz, S., Marko, B., Galle, L. 2011. The myrmecofauna (Hymenoptera: Formicidae) of Hungary: an updated checklist. North-Western Journal of Zoology 7: 55-62.
- Curtis, J. 1854. On the genus Myrmica and other indigenous ants. Trans. Linn. Soc. Lond. 21: 211-220 (page 215, Senior synonym of modesta)
- Czechowski, W. 2004. Scarcity of sites suitable for nesting promotes plesiobiosis in ants (Hymenoptera: Formicidae). Entomologica Fennica 15(4). 211–218.
- Czechowski, W., Radchenko, A., Czechowska, W. 2002. The ants (Hymenoptera, Formicidae) of Poland. MIZ PAS Warsaw.
- Dalla Torre, K. W. von. 1893. Catalogus Hymenopterorum hucusque descriptorum systematicus et synonymicus. Vol. 7. Formicidae (Heterogyna). Leipzig: W. Engelmann, 289 pp. (page 132, Senior synonym of fusca)
- Davis, T. 2009. The ants of South Carolina (thesis, Clemson University).
- Dekoninck, W., Ignace, D., Vankerkhoven, F., Wegnez, P. 2012. Verspreidingsatlas van de mieren van België. Bulletin de la Société royale belge d’Entomologie 148: 95-186.
- Devenish, A.J.M., Newton, R.J., Bridle, J.R., Gomez, C., Midgley, J.J., Sumner, S. 2021. Contrasting responses of native ant communities to invasion by an ant invader, Linepithema humile. Biological Invasions 23, 2553–2571 (doi:10.1007/s10530-021-02522-7).
- Dubovikoff, D.A., Yusupov, Z.M. 2017. Family Formicidae - Ants. In Belokobylskij S. A. and A. S. Lelej: Annotated catalogue of the Hymenoptera of Russia. Proceedingss of the Zoological Institute of the Russian Academy of Sciences 6: 197-210.
- Emery, C. 1909f. Beiträge zur Monographie der Formiciden des paläarktischen Faunengebietes. (Hym.) Teil IX. Dtsch. Entomol. Z. 1909: 695-712 (page 697, see also)
- Emery, C. 1925a [1924]. Notes critiques de myrmécologie. Ann. Soc. Entomol. Belg. 64: 177-191 (page 177, see also)
- Espadaler, X., Santamaria, S. 2012. Ecto- and Endoparasitic Fungi on Ants from the Holarctic Region. Psyche Article ID 168478, 10 pages (doi:10.1155/2012/168478).
- Fernández-Escudero, I., Tinaut, A. 1998. Heat-cold dialectic in the activity of Proformica longiseta, a thermophilous ant inhabiting a high mountain (Sierra Nevada, Spain). International Journal of Biometeorology 41, 175–182 (doi:10.1007/s004840050072).
- García, F., Cuesta-Segura, A.D., Espadaler, X. 2024. Myrmica babiensis sp. nov. (Hymenoptera: Formicidae), a new social parasite from the NW Iberian Peninsula. Annales Zoologici 74(1), 113-127 (doi:10.3161/00034541anz2024.74.1.006).
- Giannetti, D., Schifani, E., Castracani, C., Ghizzoni, M., Delaiti, M., Pfenner, F., Spotti, F.A., Mori, A., Ioriatti, C., Grasso, D.A. 2021. Assessing ant diversity in agroecosystems: the case of Italian vineyards of the Adige Valley. Redia 104, 97–109 (doi:10.19263/redia-104.21.11).
- Glaser, F. 2016. Artenspektrum, Habitatbindung und naturschutzfachliche Bedeutung von Ameisen (Hymenoptera, Formicidae) am Stutzberg (Vorarlberg, Österreich). inatura – Forschung 34: 26 S.
- Gösswald, K. 1938. Über bisher unbekannte, durch den Parasitismus der Mermithiden (Nemat.) verursachte Formveränderungen bei Ameisen. Parasitology Research 10: 138-152.
- Haelewaters, D., Boer, P., Noordijk, J. 2015. Studies of Laboulbeniales (Fungi, Ascomycota) on Myrmica ants: Rickia wasmanniii in the Netherlands. Journal of Hymenoptera Research 44, 39–47 (doi:10.3897/jhr.44.4951).
- Hashimoto, Y. 1990. Unique features of sensilla on the antennae of Formicidae (Hymenoptera). Applied Entomology and Zoology 25: 491-501.
- Hauschteck, E. 1961. Die Chromosomen von fünf Ameisenarten. Rev. Suisse Zool. 68: 218-223 (page 221, karyotype described)
- Hoey-Chamberlain, R.V. 2012. Food preference, survivorship, and intraspecific interactions of Velvety Tree Ants. M.S. thesis, University of California, Riverside.
- Imai, H. T. 1966b. The chromosome observation techniques of ants and the chromosomes of Formicinae and Myrmicinae. Acta Hymenopterol. 2: 119-131 (page 119, karyotype described)
- Ivanov, K. 2019. The ants of Ohio (Hymenoptera, Formicidae): an updated checklist. Journal of Hymenoptera Research 70: 65–87 (doi:10.3897@jhr.70.35207).
- Jansen, G., Savolainen, R. 2010. Molecular phylogeny of the ant tribe Myrmicini (Hymenoptera: Formicidae). Zoological Journal of the Linnean Society 160(3), 482–495 (doi:10.1111/j.1096-3642.2009.00604.x).
- Kanizsai, O., Lőrinczi, G., Gallé, L. 2013. Nesting associations without interdependence: A preliminary review on plesiobiosis in ants. Psyche 2013, 238602 (doi:10.1155/2013/238602).
- Kiran, K., Karaman, C. 2020. Additions to the ant fauna of Turkey (Hymenoptera, Formicidae). Zoosystema 42(18), 285-329 (doi:10.5252/zoosystema2020v42a18).
- Kiran, K., Karaman, C., Lapeva-Gjonova, A., Aksoy, V. 2017. Two new species of the "ultimate" parasitic ant genus Teleutomyrmex KUTTER, 1950 (Hymenoptera: Formicidae) from the Western Palaearctic. Myrmecological News 25, 145-155.
- Klarica, J.; Bittner, L.; Pallua, J.; Pezzei, C.; Huck-Pezzei, V.; Dowell, F.; Schied, J.; Bonn, G. K.; Huck, C.; Schlick-Steiner, B. C.; Steiner, F. M. 2011. Near-infrared imaging spectroscopy as a tool to discriminate two cryptic Tetramorium ant species. Journal of Chemical Ecology 37:549-552.
- Kök, Ş., Aktaç, N., Kasap, I. 2021. Ant (Hymenoptera: Formicidae) ‐ aphid (Hemiptera: Aphididae) interactions in different habitats from Turkey with new mutualistic associations. Agricultural and Forest Entomology 12477 (doi:10.1111/afe.12477).
- Kupyanskaya, A. N. 1990a. Ants of the Far Eastern USSR. Vladivostok: Akademiya Nauk SSSR, 258 pp. (page 151, see also)
- Kutter, H. 1958. Über Modificationen bei Ameisenarbeiterinnen, welche durch den Parasitismus von Mermithiden (Nematod.) verursacht worden sind. Mitteilungen der Schweizerischen Entomologischen Gesellschaft 31: 313-316.
- Kutter, H. 1977c. Hymenoptera, Formicidae. Insecta Helv. Fauna 6: 1-298 (page 157, see also)
- Kwon, T.-S. 2015. Ant assemblages along the Baekdudaegan Mountain Range in South Korea: Human roads and temperature niche. Journal of Asia-Pacific Biodiversity 8, 152–157 (doi:10.1016/j.japb.2015.05.001).
- Laciny, A. 2021. Among the shapeshifters: parasite-induced morphologies in ants (Hymenoptera, Formicidae) and their relevance within the EcoEvoDevo framework. EvoDevo 12, 2 (doi:10.1186/s13227-021-00173-2).
- Lamelas-Lopez, L., Gabriel, R., Ros-Prieto, A., Borges, P. 2023. SLAM Project - Long Term Ecological Study of the Impacts of Climate Change in the natural forest of Azores: VI - Inventory of Arthropods of Azorean Urban Gardens. Biodiversity Data Journal 11, e98286 (doi:10.3897/bdj.11.e98286).
- Lapeva-Gjonova, A., Antonova, V. 2022. An updated checklist of ants (Hymenoptera, Formicidae) of Bulgaria, after 130 years of research. Biodiversity Data Journal 10, e95599 (doi:10.3897/bdj.10.e95599).
- Lapeva-Gjonova, A., Antonova, V., Ljubomirov, T. 2021. Ants (Hymenoptera, Formicidae) of Sarnena Sredna Gora Mountains (Bulgaria). Fauna of Sarnena Sredna Gora Mts, Part 2 ZooNotes, Supplement 10: 18-27.
- Lapeva-Gjonova, A., Radchenko, A. 2021. Ant genus Strongylognathus (Hymenoptera, Formicidae) in Bulgaria: a preliminary review. Biodiversity Data Journal 9, e65742 (doi:10.3897/bdj.9.e65742).
- Latreille, P. A. 1798. Essai sur l'histoire des fourmis de la France. Brive: F. Bourdeaux, 50 pp. (page 50, queen, male described)
- Lee, C.-C., Weng, Y.-M., Lai, L.-C., Suarez, A.V., Wu, W.-J., Lin, C.-C., Yang, C.-C.S. 2020. Analysis of recent interception records reveals frequent transport of arboreal ants and potential predictors for ant invasion in Taiwan. Insects 11, 356 (doi:10.3390/INSECTS11060356).
- Linnaeus, C. 1758. Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus I. Editio decima, reformata. Holmiae [= Stockholm]: L. Salvii, 824 pp. (page 581, worker described)
- López, F. 1991a. Estudio morfológico y taxonómico de los grupos de especies ibéricas del género Tetramorium Mayr, 1855 (Hym., Formicidae). Bol. Asoc. Esp. Entomol. 15: 29-52 (page 31, see also)
- López, F. 1991b. Variabilidad morfológica y problemas taxonómicos en Tetramorium caespitum (Linné, 1758) y Tetramorium semilaeve André, 1881 (Hym., Formicidae). Bol. Asoc. Esp. Entomol. 15: 65-78 (page 73, see also)
- Mackay, W. P. and E. Mackay. 2002. The ants of New Mexico (Hymenoptera: Formicidae). Edwin Mellen Press, Lewiston, NY.
- Mayr, G. 1855. Formicina austriaca. Beschreibung der bisher im österreichischen Kaiserstaate aufgefundenen Ameisen, nebst Hinzufügung jener in Deutschland, in der Schweiz und in Italien vorkommenden Arten. Verh. Zool.-Bot. Ver. Wien 5: 273-478 (page 426, Combination in Tetramorium, Senior synonym of modesta)
- Mayr, G. 1861. Die europäischen Formiciden. Nach der analytischen Methode bearbeitet. Wien: C. Gerolds Sohn, 80 pp. (page 62, queen, male described)
- Moss, A.D., Swallow, J.G., Greene, M.J. 2022. Always under foot: Tetramorium immigrans (Hymenoptera: Formicidae), a review. Myrmecological News 32: 75-92 (doi:10.25849/MYRMECOL.NEWS_032:075).
- Moura, M.N., Cardoso, D.C., Cristiano, M.P. 2020. The tight genome size of ants: diversity and evolution under ancestral state reconstruction and base composition. Zoological Journal of the Linnean Society, zlaa135 (doi:10.1093/zoolinnean/zlaa135).
- Moya-Raygoza, G., Martinez, A.V. 2014. Ants (Hymenoptera: Formicidae) and trophobiont leafhopper nymphs (Hemiptera: Cicadellidae) become more abundant in shaded donditions: Implications for mutualism. Florida Entomologist 97, 1378–1385 (doi:10.1653/024.097.0412).
- Nemet, E., Czekes, Z., Tausan, I., Marko, B. 2012. Contribution to the knowledge of the myrmecofauna of the Cefa Nature Park (North-Western Romania). Acta Scientiarum Transylvanica Biologia 20, 61-72.
- Orou, N., Csősz, S., Arnan, X., Pol, R. G., Arthofer, W., Schlick-Steiner, B. C., Steiner, F. M. 2023. Messor erwini sp. n., a hitherto cryptic harvester ant in the Iberian Peninsula. Zoologischer Anzeiger 307, 36–53 (doi:10.1016/j.jcz.2023.09.001).
- Pashaei Rad, S., Taylor, B., Torabi, R., Aram, E., Abolfathi, G., Afshari, R., Borjali, F., Ghatei, M., Hediary, F., Jazini, F., Heidary Kiah, V., Mahmoudi, Z., Safariyan, F., Seiri, M. 2018. Further records of ants (Hymenoptera: Formicidae) from Iran. Zoology in the Middle East 64, 145-159 (doi:10.1080/09397140.2018.1442301).
- Pekár, S., Haddad, C. 2011. Trophic strategy of ant-eating Mexcala elegans (Araneae: Salticidae): looking for evidence of evolution of prey-specialization. Journal of Arachnology 39, 133–138 (doi:10.1636/hi10-56.1).
- Pérez‐Marcos, M., López‐Gallego, E., Arnaldos, M.I., Martínez‐Ibáñez, D., García, M.D. 2020. Formicidae (Hymenoptera) community in corpses at different altitudes in a semiarid wild environment in the southeast of the Iberian Peninsula. Entomological Science 23, 297–310 (doi:10.1111/ENS.12422).
- Purkart, A., Kollár, J., Goffová, K. 2019. Fauna of ants (Hymenoptera: Formicidae) of selected sand habitats in Podunajsko Region, Slovakia. Naturae Tutela 23(1): 101-111.
- Purkart, A., Wagner, H.C., Goffová, K., Selnekovič, D., Holecová, M. 2021. Laboratory observations on Anergates atratulus (Schenck, 1852): mating behaviour, incorporation into host colonies, and competition with Strongylognathus testaceus (Schenck, 1852). Biologia (doi:10.1007/s11756-021-00901-y).
- Radchenko, A. G. 1992b. Ants of the genus Tetramorium (Hymenoptera, Formicidae) of the USSR fauna. Report 2. Zool. Zh. 71(8 8: 50-58 (page 50, Senior synonym of himalayanum, indocile and transbaicalense)
- Radchenko, A. G.; Czechowski, W.; Czechowska, W. 1998. The genus Tetramorium Mayr (Hymenoptera, Formicidae) in Poland - a survey of species and a key for their identification. Ann. Zool. (Warsaw) 48: 107-118 (page 108, see also)
- Radchenko, A.G., Fisher, B.L., Esteves, F.A., Martynova, E.V., Bazhenova, T.N., Lasarenko, S.N. 2023. Ant type specimens (Hymenoptera, Formicidae) in the collection of Volodymyr Opanasovych Karawajew. Communication 1. Dorylinae, Poneromorpha and Pseudomyrmecinae. Zootaxa, 5244(1), 1–32 (doi:10.11646/zootaxa.5244.1.1).
- Rahiminejad, V., Hajiqanbar, H. 2020. New records of mites of the Heterostigmata (Acari: Prostigmata) associated with insects from Golestan Province, northern Iran. Persian Journal of Acarology 9(3): 233–242 (doi:10.22073/pja.v9i3.61223).
- Rasheed, M.T., Bodlah, I., Fareen, A.G., Wachkoo, A.A., Huang, X., Akbar, S.A. 2019. A checklist of ants (Hymenoptera: Formicidae) in Pakistan. Sociobiology 66(3), 426-439 (doi:10.13102/sociobiology.v66i3.4330).
- Rericha, L. 2007. Ants of Indiana. Indiana Department of Natural Resources, 51pp.
- Reznikova, Z. 2020. Spatial cognition in the context of foraging styles and information transfer in ants. Animal Cognition. (doi:10.1007/s10071-020-01423-x).
- Rigato, F.; Toni, I. 2011. Short notes 21. Hymenoptera, Formicidae. Pp. 873-882 in: Nardi, G.; Whitmore, D.; Bardiani, M.; Birtele, D.; Mason, F.; Spada, L.; Cerretti, P. (eds.) 2011. Biodiversity of Marganai and Montimannu (Sardinia). Research in the framework of the ICP Forests network. Conservazione Habitat Invertebrati, 5. Sommacampagna, Verona: Cierre Edizioni, 896 pp.
- Ruano, F., Tinaut, A., Soler, J.J. 2000. High surface temperatures select for individual foraging in ants. Behavioral Ecology 11, 396-404.
- Ryu, J., Kim, Y.-K., Suh, S.J., Choi, K.S. 2021. The Insect database in Dokdo, Korea: An updated version in 2020. Biodiversity Data Journal 9, e62011 (doi:10.3897/bdj.9.e62011).
- Salata, S., Borowiec, L. 2019. Comments to distribution of several Greek Tetramorium Mayr, 1855 species (Hymenoptera: Formicidae). Annals of the Upper Silesian Museum in Bytom, Entomology 28, 1-9 (doi:10.5281/ZENODO.2644897).
- Salata, S., Borowiec, L., Trichas, A. 2020. Review of ants (Hymenoptera: Formicidae) of Crete, with keys to species determination and zoogeographical remarks. Monographs of the Upper Silesian Museum No 12: 5–296 (doi:10.5281/ZENODO.3738001).
- Salata, S., Khalili-Moghadam, A., Borowiec, L. 2024. A new species of the Tetramorium meridionale species-group (Hymenoptera: Formicidae) from Iran. Zoology in the Middle East 70, 161–175 (doi:10.1080/09397140.2024.2359167).
- Salata, S., van Delft, J.P.L., van Delft, J.J.C.W., Georgiadis, C., Borowiec, L. 2023. Tetramorium albenae Salata, van Delft & Borowiec n. sp. (Hymenoptera, Formicidae) – a new inquiline ant species from the Balkan Peninsula. The European Zoological Journal 90(1), 333–343 (doi:10.1080/24750263.2023.2198548).
- Sanetra, M., Buschinger, A. 2000. Phylogenetic relationships among social parasites and their hosts in the ant tribe Tetramoriini (Hymenoptera: Formicidae). European Journal of Entomology 97: 95-117.
- Sanetra, M., Güsten, R., Schulz, A. 1999. On the taxonomy and distribution of Italian Tetramorium species and their social parasites (Hymenoptera Formicidae). Memorie della Societa Entomologica Italiana 77: 317-357.
- Schifani, E., Csősz, S., Viviano, R., Alicata, A. 2021. Ant diversity on the largest Mediterranean islands: on the presence or absence of 28 species in Sicily (Hymenoptera, Formicidae). Natural History Sciences 8, 55–70 (doi:10.4081/nhs.2021.532).
- Schifani, E., Nalini, E., Gentile, V., Alamanni, F., Ancona, C., Caria, M., Cillo, D., Bazzato, E. 2021. Ants of Sardinia: An updated checklist based on new faunistic, morphological and biogeographical notes. Redia 104, 21–35 (doi:10.19263/redia-104.21.03).
- Schultner, E., Pulliainen, U. 2020. Brood recognition and discrimination in ants. Insectes Sociaux 67, 11–34 (doi:10.1007/s00040-019-00747-3).
- Seifert, B. 2018. The Ants of Central and North Europe. lutra Verlags– und Vertriebsgesellschaft, Tauer, Germany, 408 pp.
- Seifert, B. 2021. Surviving the winter: Tetramorium sibiricum n. sp., a new Central Siberian ant species (Hymenoptera: Formicidae). Osmia 9, 15–24 (doi:10.47446/osmia9.3).
- Sharaf, M.R., Abdel-Dayem, M.S., Mohamed, A.A., Fisher, B.L., Aldawood, A.S. 2020. A preliminary synopsis of the ant fauna (Hymenoptera: Formicidae) of Qatar with remarks on the zoogeography. Annales Zoologici 70: 533-560 (doi:10.3161/00034541anz2020.70.4.005).
- Shiran, E., Mossadegh, M.S., Esfandiari, M. 2013. Mutualistic ants (Hymenoptera: Formicidae) associated with aphids in central and southwestern parts of Iran. Journal of Crop Protection 2: 1-12.
- Siddiqui, J. A., Li, J., Zou, X., Bodlah, I., Huang, X. 2019. Meta-analysis of the global diversity and spatial patterns of aphid-ant mutualistic relationships. Applied Ecology and Environmental Research 17: 5471-5524 (doi:10.15666/aeer/1703_54715524).
- Smith, D. R. 1979. Superfamily Formicoidea. Pp. 1323-1467 in: Krombein, K. V., Hurd, P. D., Smith, D. R., Burks, B. D. (eds.) Catalog of Hymenoptera in America north of Mexico. Volume 2. Apocrita (Aculeata). Washington, D.C.: Smithsonian Institution Pr (page 1400, see also)
- Smith, F. 1851. List of the specimens of British animals in the collection of the British Museum. Part VI. Hymenoptera, Aculeata. London: British Museum, 134 pp. (page 118, Senior synonym of fuscula)
- Snegovaya, N., Shigayev, C. 2021. A checklist of the ants (Insecta, Formicidae) of Azerbaijan Republic. Iranian Journal of Animal Biosystematics 17(2): 179-207 (doi:10.22067/ijab.2022.67343.1000).
- Sondej, I., Domisch, T. 2024. Impact of large-scale fire and habitat type on ant nest density and species abundance in Biebrza National Park, Poland. Forests 151, 123 (doi:10.3390/f15010123).
- Stankovic, B. 2021. Contribution to the knowledge of the myrmecofauna (Hymenoptera: Formicidae) of Pomorie, Bulgaria. Ahi Evran International Conference on Scientific Research, pp. 198-204.
- Stukalyuk, S., Radchenko, A., Akhmedov, A., Reshetov, A., Netsvetov, M. 2021. Acquisition of invasive traits in ant, Crematogaster subdentata Mayr (Hymenoptera: Formicidae) in urban environments. Serangga 26: 1-29.
- Stukalyuk, S.V., Radchenko, A., Reshetov, A., Akhmedov, A., Goncharenko, I. 2021. Comparative analysis of the population structure of Crematogaster subdentata and Lasius neglectus in the primary and secondary ranges (Hymenoptera: Formicidae). Fragmenta Entomologica 53, 43-51 (doi:10.13133/2284-4880/436).
- Wagner, H.C., Arthofer, W., Seifert, B., Muster, C., Steiner, F.M. & Schlick-Steiner, B.C. 2017. Light at the end of the tunnel: Integrative taxonomy delimits cryptic species in the Tetramorium caespitum complex. Myrmecological News 25: 95-129.
- Wahab, A. 1962. Untersuchungen uber Nematoden in den drusen des kopfes der Ameisen (Formicidae). Zeitschrift fur Morphologie und Okologie der Tiere 52: 33–92.
- Waters, J.S., Keough, N.W., Burt, J., Eckel, J.D., Hutchinson, T., Ewanchuk, J., Rock, M., Markert, J.A., Axen, H.J., Gregg, D. 2022. Survey of ants (Hymenoptera, Formicidae) in the city of Providence (Rhode Island, United States) and a new northern-most record for Brachyponera chinensis (Emery, 1895). Check List 18(6), 1347–1368 (doi:10.15560/18.6.1347).
- Wegnez, P. 2017. Découverte de Myrmica lobicornis Nylander, 1846 et Lasius jensi Seifert, 1982, deux nouvelles espèces pour le Grand-Duché de Luxembourg (Hymenoptera: Formicidae). Bulletin de la Société royale belge d’Entomologie153, 46–49.
- Wetterer, J.K., Wetterer, A.L. 2004. Ants (Hymenoptera: Formicidae) of Bermuda. Florida Entomologist 87(2), 212–221 (doi:10.1653/0015-4040(2004)087[0212:ahfob2.0.CO;2]).
- Wheeler, G. C. and J. Wheeler. 1986. The ants of Nevada. Natural History Museum of Los Angeles County, Los Angeles.
- Wheeler, G. C.; Wheeler, J. 1954d. The ant larvae of the myrmicine tribes Meranoplini, Ochetomyrmicini and Tetramoriini. Am. Midl. Nat. 52: 443-452 (page 445, larva described)
- Wiezik, M., Svitok, M., Wieziková, A., Dovčiak, M. 2013. Shrub encroachment alters composition and diversity of ant communities in abandoned grasslands of western Carpathians. Biodiversity and Conservation 22, 2305–2320 (doi:10.1007/s10531-013-0446-z).
- Wu, J. & Wang, C. 1992. Formicidae (pp. 1301-1320). In Peng, J. et al. Iconography of Forest Insects in Hunan, China. Forest Bureau of Hunan Province: 1473 pp. Hunan Scientific and Technical Publishing House.
- Wu, J., Wang, C. 1995. The ants of China. Beijing: China Forestry Publishing House, x + 214 pp.
- Yamane, S., Hosoishi, S., Ito, F. 2022. Japanese Tetramorium queens: identification key and species diagnoses (Hymenoptera, Formicidae, Myrmicinae). ZooKeys 1084: 43–64 (doi:10.3897/zookeys.1084.69767).
References based on Global Ant Biodiversity Informatics
- Abe T. 1971. On the food sharing among four species of ants in a sandy grassland. I. Food and foraging behaviour. Japanese Journal of Ecology 20(6): 219-230.
- Abril S., C. Gómez. 2012. Lista actualizada de las especies de hormigas de Menorca (Islas Baleares, España) y primera cita de Monomorium andrei Saunders, 1890 (Hymenoptera, Formicidae) de la isla Boletín de la Sociedad Entomológica Aragonesa 50: 403-407.
- Agosti, D. and C.A. Collingwood. 1987. A provisional list of the Balkan ants (Hym. Formicidae) and a key to the worker caste. I. Synonymic list. Mitteilungen der Schweizerischen Entomologischen Gesellschaft, 60: 51-62
- Aktaç, N. "Studies on the myrmecofauna of Turkey I. Ants of Siirt, Bodrum and Trabzon." Istanbul Universitesi Fen Fakultesi Mecmuasi. Seri B 41 (1977): 115-135.
- Aldawood AS, Sharaf MR (2011) Monomorium dryhimi sp. n., a new ant species (Hymenoptera, Formicidae) of the M. monomorium group from Saudi Arabia, with a key to the Arabian Monomorium monomorium-group. ZooKeys 106: 4754. doi: 10.3897/zookeys.106.139
- Alvarado M., and L. Galle. 2000. Ant assemblages associated with lowland forests in the southern part of the great Hungarian plain. Acta Zoologica Academiae Scientarum Hungaricae 46(2): 79-102.
- Andoni V. 1977. Kontribut mbi Himenopteret e familjes Formicidae te vendit tone. Buletini I Shkencave te Natyres 31(2): 93-101.
- Anonymous 1986. A list of ants collected at Akiyoshi-dai (Yamaguchi-ken) by members of the Myrmecologists Society (Japan) in 1985. Ari 14:5-6.
- Anonymous. A list of ants collected at Bandai, Fukushima Prefecture by the members of the Myrmecological Society of Japan in 1992. ARI Reports of the Myrmecologists Society (Japan) 18: 31
- AntArea. Accessed on February 5th 2014 at http://antarea.fr/fourmi/
- Antonov I. A. 2013. Ant Assemblages (Hymenoptera: Formicidae) of Cities of the Temperate Zone of Eurasia. Russian Journal of Ecology 44(6): 523526.
- Arnan X., M. Gracia, L. Comas, and J. Retana. 2009. Forest management conditioning ground ant community structure and composition in temperate conifer forests in the Pyrenees Mountains. Forest Ecology and Management 258(2): 5159.
- Arnan, X., A. Rodrigo and J. Retana. 2006. Post-fire recovery of Mediterranean ground ant communities follows vegetation and dryness gradients. Journal of Biogeography 33(7):1246-1258
- ArtDatabanken Bugs (via GBIG)
- Asociacion Iberica de Mirmecologia. 2012. List of species collected during the Taxomara Tres Cantos 2012. Iberomyrmex 4: 17-18.
- Asociacion Iberica de Mirmecologia. 2016. List of species collected during the Taxomara Murcia 2016. Iberomyrmex 8: 48-49.
- Asso, I. "Introductio in Oryctographiam et Zoologian Aragonae." (1784).
- Astric A., and H. Callot. 2006. Presence d'Anergates atratulus (Schenck, 1852) et Strongylognathus testaceus (Schenck, 1852) en Alsace. Bull. Soc. Ent. Mulhouse 62(2): 19-21.
- Azuma M. 1977. On the myrmecological-fauna of Mt. Rokko, Hyogo, with description of a new species (Formicidae, Hymenoptera). Hyogo Biology 7:112-118.
- Baroni Urbani C. 1968. Studi sulla mirmecofauna d'Italia. IV. La fauna mirmecologica delle isole Maltesi ed il suo significato ecologico e biogeografico. Ann. Mus. Civ. Stor. Nat. Giacomo Doria 77: 408-559.
- Baroni Urbani C. 1968. Studi sulla mirmecofauna d'Italia. V. Aspetti ecologici della Riviera del M. Cònero. Boll. Zool. 35: 39-76.
- Baroni Urbani C., and C. A. Collingwood. 1976. A Numerical Analysis of the Distribution of British Formicidae (Hymenoptera, Aculeata). Verhandlungen der Naturforschenden Gesellschaft in Basel 85: 51-91.
- Baroni Urbani C., and C. A. Collingwood. 1977. The zoogeography of ants (Hymenoptera, Formicidae) in Northern Europe. Acta Zoologica Fennica 152: 1-34.
- Baroni Urbani, C. "Formiche dell'Italia appenninica (Studi sulla mirmecofauna d'Italia, III)." Memorie del Museo Civico di Storia Naturale di Verona 12 (1964): 149-172.
- Baroni Urbani, C. "Studi sulla mirmecofauna d'Italia. II. Formiche di Sicilia." Atti dell'Accademia Gioenia di Scienze Naturali in Catania (6) 16 (1964): 25-66.
- Baroni Urbani, C. "Su alcune formiche raccolte in Turchia." Annuario dell'Istituto e Museo di Zoologia dell'Università di Napoli 16 (1964): 1-12.
- Baroni-Urbani 1969. Ant Communities of the High-Altitude Appennine Grasslands. Ecology 50(3): 488-492.
- Barrett K. E. 1967. Ants in South Brittany. Entomologist's Record and Journal of Variation 79:112-116.
- Barrett K. E. J. 1968. Ants in western France. Entomologist 101: 153-155.
- Barrett K. E. J. 1968b. The distribution of ants in central southern England. Transactions of the Society for British Entomology 17: 235-250.
- Barrett K. E. J. 1970. Ants in France, 1968-69. Entomologist 103: 270-274.
- Baugnee J. Y., and J. F. Godeau. 2000. Signalement de Monomorium monomorium Bolton et de quelques autres fourmis à Belle-Ile-en-Mer, en Bretagne (Hymenoptera, Formicidae). Bulletin de la Société entomologique de France 105(2): 205-208.
- Bernard F. 1958. Les fourmis des iles Pelagie, comparaison avec d'autres faunes insulaires. Esttrato dalla Rivista di Biologia Coloniale XVI: 67-79.
- Bernard F. 1959. Les fourmis de l'île de Port-Cros. Contribution à l'écologie des anciennes forêts méditerranéennes. Vie Milieu 9: 340-360.
- Bernard F. 1960. Fourmis récoltées en Corse par J. Bonfils (1957). Compte Rendu Sommaire des Séances de la Société de Biogéographie 36: 108-114.
- Bernard F. 1967. Recherches sur les fourmis des Monts-Dore. Annales de la Station biologique de Besse-en-Chandesse : 1-11.
- Bernard F. 1973. Etudes ecologiques sur les fourmis de Breuil-Cervinia (Val d"Aoste). Revue Ecol Biol Sol, 102: 237-269.
- Bernard F. 1975. Rapports entre fourmis et vegetation pres des Gorges du Verdon. Annales du Muséum d'Histoire Naturelle de Nice 2: 57-79.
- Bernard F. 1977. Écologie des fourmis du Parc national de Port-Cros. Bulletin du Muséum National d'Histoire Naturelle. Écologie Générale (3)36: 53-82.
- Bernard F. 1978. Fourmis et milieu dans le massif des Maures. Vie et Milieu. Série C. Biologie Terrestre 27: 83-118.
- Bernard, F. "Notes sur les fourmis de France. II. Peuplement des montagnes méridionales." Annales de la Société Entomologique de France 115 (1950): 1-36.
- Bernard, F. "Remarques sur le peuplement des Baléares en fourmis." Bulletin de la Société d' Histoire naturelle de l' Afrique du Nord 47 (1956): 254-266.
- Bezdeckova K., and P. Bezdecka. 2008. The ants of the Bohemian-Moravian Highlands. Acta rerum naturalium 5: 243252.
- Bigot L. 1959. Complement a l'inventaire de la faune entomologique de la Camargue 93eme note). La Terre et la Vie 106 : 149-157.
- Blacker N. C. 1989. The ants (Hymenoptera, Formicidae) of the Gower Peninsula, West Glamorgan, South Wales. Entomologist's Record and Journal of Variation 101: 261-266.
- Blacker N.C. 2007. Ants (Hym., Formicidae) in East Anglia-Additional Records from . Entomologist's Monthly Magazine 143: 69-90
- Blinova S. V. 2011. Changes in the Ant Assemblage of PineBirch Forest upon Removal of the Nests of Dominant Species. Russian Journal of Ecology 42(6): 525528.
- Boer P. 2019. Species list of the Netherlands. Accessed on January 22 2019 at http://www.nlmieren.nl/websitepages/specieslist.html
- Boer P., W. Dekoninck, A. J. van Loon, and F. Vankerkhoven. 2003. Lijst van mieren (Hymenoptera: Formicidae) van Belgie en Nederland, hun Nederlandse namen en hun voorkomen. Entomologische Berichten 63(3): 54-57.
- Boer P., W. Dekoninck, A. J. van Loon, and F. Vankerkhoven. 2003. List of ants (Hymenoptera: Formicidae) of Belgium and The Netherlands, their status and Dutch vernacular names. Entomologische Berichten 63 (3): 54-58.
- Boer P., and J. Noordijk. 2004. De ruige gaststeekmier Myrmica hirsuta nieuw voor Nederland (Hymenoptera: Formicidae). Ned. Faun. Meded. 20: 25-32.
- Bonaric J. C. 1971. Contribution a l'etude systematique et ecologique des formicides du Bas-Languedoc. PhD thesis Universite des sciences et techniques du Languedoc, 175 pages.
- Bonaric J. C. 1971. Étude systématique et ecologique des fourmis de lHérault (suite). Ann. Soc. Hortic. Hist. Nat. Hérault 111: 119-126.
- Bonte D., W. Dekoninck, S. Provoost, E. Cosijns, and M. Hoffmann. 2003. Microgeographical distribution of ants (Hymenoptera: Formicidae) in coastal dune grassland and their relation to the soil structure and vegetation. Animal Biology 53(4): 367-377.
- Borowiec L. 2014. Catalogue of ants of Europe, the Mediterranean Basin and adjacent regions (Hymenoptera: Formicidae). Genus (Wroclaw) 25(1-2): 1-340.
- Borowiec L., and S. Salata. 2012. Ants of Greece - Checklist, comments and new faunistic data (Hymenoptera: Formicidae). Genus 23(4): 461-563.
- Bracko, G. 2006. Review of the ant fauna (Hymenoptera:Formicidae) of Croatia. Acta Entomologica Slovenica 14(2): 131-156.
- Bracko, G. "Review of the ant fauna (Hymenoptera: Formicidae) of Croatia." Acta Entomologica Slovenica Vol 14 st (2006): 131-156.
- Braschler, B. and B. Baur. 2005. Experimental Small-Scale Grassland Fragmentation Alters Competitive Interactions among Ant Species. Oecologia 143(2):291-300
- Buschinger, A., W. Ehrhardt, and K. Fischer. "Doronomyrmex pacis, Epimyrma stumperi und E. goesswaldi (Hym., Formicidae) neu für Frankreich." Insectes Sociaux 28 (1981): 67-70.
- Cagniant H. 1997. Le genre Tetramorium au Maroc (Hymenoptera: Formicidae): clé et catalogue des espèces. Ann. Soc. Entomol. Fr. (N.S.) 33: 89-100.
- Cagniant, H. 2006. Liste actualisée des fourmis du Maroc. Myrmecologische Nachrichten 8: 193-200.
- Cagniant, H., and X. Espadaler. "Liste des espèces de fourmis du Maroc." Actes des Colloques Insectes Sociaux 8 (1993): 89-93.
- Cagniant, H. "Liste actualisee des fourmis du Maroc (Hymenoptera: Formicidae)." Myrmecologische Nachrichten 8 (2006): 193-200.
- Cagniant, H. "Étude de fourmis récoltées par le Professeur H. Janetschek dans la Sierra Nevada." Bulletin de la Société d' Histoire naturelle de l' Afrique du Nord 52 (1961): 104-117.
- Cammell, M. E., M. J. Way, and M. R. Paiva. "Diversity and structure of ant communities associated with oak, pine, eucalyptus and arable habitats in Portugal." Insectes Sociaux 43 (1996): 37-46.
- Carniel A. 1998. Ricerche sulla mirmecofauna delle Prealpi Orobiche (Lombardia) (Insecta, Hymenoptera, Formicidae). Atti. Mus. Civ. Stor. Nat. Morbegno 9: 29-39.
- Carpintero S., J. Reyes-Lopes, and G. M. Luque. 2011. Ant community structure under Retama sphaerocarpa shrubs in a semi-arid environment. Entomological Science 14: 147153.
- Casevitz-Weulersse J. 1990. Etude Systematique de la Myrmecofaune Corse (Hymenoptera: Formicidae), deuxieme partie. Bull. Mus. Natn. Hist. Nat. Paris. 4eme serie 12, section A(2): 415-442.
- Casevitz-Weulersse J., and C. Galkowski. 2009. Liste actualisee des Fourmis de France (Hymenoptera, Formicidae). Bull. Soc. Entomol. Fr. 114: 475-510.
- Casevitz-Weulersse J., and M. Prost. 1991. Fourmis de la Côte-d'Or présentes dans les collections du Muséum d'Histoire Naturelle de Dijon. Bulletin Scientifique de Bourgogne 44: 53-72.
- Casevitz-Weulersse, J. "Contribution a la connaisance des fourmis de la Corse (Hymenoptera: Formicidae)." These de Doctorat Museum Nat (1989): 379pp.
- Castracani C., D. A. Grasso, A. Fanfani, and A. Mori. 2010. The ant fauna of Castelporziano Presidential Reserve (Rome, Italy) as a model for the analysis of ant community structure in relation to environmental variation in Mediterranean ecosystems. J Insect Conserv 14: 585594.
- Cerda X., R. Palacios, and J. Retana. 2009. Ant community structure in Citrus orchards in the Mediterranean basin: impoverishment as a consequence of habitat homogeneity. Environ. Entomol. 38(2): 317-324.
- Chang Youde, and He Dahan. 2001. A taxonomic study of the ant genus Tetramorium Mayr (Hymenoptera; Formicidae) in Northwest China. Journal of Ningxia Agricultural College 22(1): 1-7.
- Cherix D., and S. Higashi. 1979. Distribution verticale des fourmis dans le Jura vaudois et recensement prelimaire des bourdons (Hymenoptera, Formicidae et Apidae). Bull. Soc. Vaud. Sc. Nat. 356(74): 315-324.
- Choe L. J., K. J. Cho, S. K. Choi, S. H. Lee, M. K. Kim, H. S. Bang, J. Eo, and M. H. Kim. 2016. Effects of landscape and management on ground-dwelling insect assemblages of farmland in Jeju Island, Korea. Entomological Research 46: 36–44.
- Choi B.-M. 1987. Taxonomic study on ants (Formicidae) in Korea (1). On the genus Monomorium. Journal of the Institute of Science Education (Cheongju National Teachers' College) 11:17-30.
- Choi B.M. 1986. Studies on the distribution of ants (Formicidae) in Korea. Journal of Chongju National Teacher College 23: 317-386.
- Choi B.M. 1988. Studies on the distribution of ants (Formicidae) in Korea (5) Ant fauna in Is. Kanghwado. Chongju Sabom Taehak Nonmunjip (Journal of Chongju National Teacher' College) 25: 217-231.
- Choi B.M. 1996. Distribution of ants (Formicidae) in Korea (16) - Ant fauna from Chollabukdo. Korean J. Soil. Zoology 1(1): 5-23.
- Choi B.M. 1996. Distribution of ants (Formicidae) in Korea (16): Ant fauna from Chollabukdo. Korean Journal of Soil Zoology 1(1): 5-23.
- Choi B.M. 1996. Studies on the distribution of ants (Formicidae) in Korea (15) -Ant fauna islands Ullungdo and Dokdo. Journal of Chongju National University of Education 33: 201-219.
- Choi B.M. 1997. Distribution of Ants (Formicidae) in Korea (18). Ants Fauna in island Paekryongdo and Taechongdo. Journal of Chongju National University of Education 34: 119-138.
- Choi B.M. 1998. Distribution of ants (Formicidae) in Korea (19) Ants fauna from Chungcheongbukdo Province. Cheongju Tea. Coll. 35: 213-266.
- Choi B.M. and C.H. Kim, 1987, Studies on the distribution of ants (Formicidae) in Korea (4). Ant fauna in Is. Hongdo and Is. Taehukusando. Journal of Chongju National Teacher College 24: 357-370.
- Choi B.M., Bang, J.R. 1992. Studies on the distribution of ants (Formicidae) in Korea (9). Ant fauna in Mt. Togyusan. Korean Journal of Applied Entomology 31:101-112.
- Choi B.M., E.C. Park. 1998. Studies on the distribution of ants (Formicidae) in Korea (20). Ant fauna in Chiaksan. Korean J. Soil Zoology 3(2): 58-62.
- Choi B.M., I. H. Lee. 1995. Studies on the distribution of ants (Formicidae) in Korea (14). Ant fauna in island Sohuksando. Korean Journal of Applied Entomology 34(3): 191-197.
- Choi B.M., K. Ogata, and M. Terayama. 1993. Comparative studies of ant faunas of Korea and Japan. 1. Faunal comparison among islands of Southern Korean and northern Kyushu, Japan. Bull. Biogeogr. Soc. Japan 48(1): 37-49.
- Choi B.M., Kim, C.H., Bang, J.R. 1993. Studies on the distribution of ants (Formicidae) in Korea (13). A checklist of ants from each province (Do), with taxonomic notes. Cheongju Sabom Taehakkyo Nonmunjip (Journal of Cheongju National University of Education) 30: 331-380.
- Choi B.M., Park K.S. 1991. Studies on the distribution of ants (Formicidae) in Korea (6). The vegetation, the species composition and the colony density of ants in Mt. Namsam, Seoul. Korean Journal of Applied Entomology 30:65-79.
- Choi B.M., and E.C. Park. 1999. Studies on the distribution of ants (Formicidae) in Korea (23) - Ant fauna Mt Huksong. Sci. Edu. Cheongju Univ. of Education 20: 21-26.
- Choi B.M., and J. R. Bang. Studies on the distribution of ants (Formicidae) in Korea (12): the analysis of ant communities in 23 islands. Journal of Cheongju National University of Education 30:317-330.
- Choi B.M., and Park, K.S. 1991. Studies on the distribution of ants (Formicidae) in Korea (7). Ant fauna in Mt. Kyeryongsan. Korean Journal of Applied Entomology 30: 80-85.
- Choi B.M.; Bang, J.R. 1992. Studies on the distribution of ants (Formicidae) in Korea (10). Ant distribution in Gangweon Do. Journal of the Institute of Science Education (Cheongju National Teachers' College) 14:12-30.
- Collingwood C. A. 1956. Ant hunting in France. Entomologist 89: 106-108.
- Collingwood C. A. 1961. Ants in the Scottish Highlands. Scotish Naturalist 70: 12-21.
- Collingwood C. A. 1971. A synopsis of the Formicidae of north Europe. Entomologist 104: 150-176
- Collingwood C. A. 1976. Ants (Hymenoptera: Formicidae) from North Korea. Annales Historico-Naturales Musei Nationalis Hungarici 68:
- Collingwood C. A. 1981. Ants (Hymenoptera: Formicidae) from Korea, 2. Folia Entomologica Hungarica 42(34): 25-30.
- Collingwood C., and A. Prince. 1998. A guide to ants of continental Portugal (Hymenoptera: Formicidae). Boletim da Sociedade Portuguesa de Entomologia. Suplemento 5: 1-49.
- Collingwood C., and H. Heatwole. 2000. Ants from Northwestern China (Hymenoptera, Formicidae). Psyche 103 (1-2): 1-24.
- Collingwood C.A. 1959. Ants in the Scottish Highlands. The Scottish Naturalist. 70: 12-21
- Collingwood C.A. 1959. Scandinavian Ants. Entomol. Rec. 71: 78-83
- Collingwood C.A. 1961. Ants in Finland. Entomol. Rec. 73: 190-195
- Collingwood C.A. 1961. New Vice-County Records for British Ants. Entomologist. 73: 90-93
- Collingwood, C. A. 1964. The Identification of British Ants (Hym. Formicidae). Transactions of the Society for British Entomology. 16:93-121.
- Collingwood, C. A. 1974. A revised list of Norwegian ants (Hymenoptera: Formicidae). Norsk Entomologisk Tidsskrift 21: 31-35.
- Collingwood, C. A. 1993. A Comparitive Study of the Ant Fauna of Five Greek Islands. Biologia Gallo-hellenica. 20,1:191-197
- Collingwood, C. A., and I. H. H. Yarrow. "A survey of Iberian Formicidae." EOS (Revista española de entomología) 44 (1969): 53-101.
- Collingwood, C. A. "A comparative study of the ant fauna of five Greek islands." Biologia Gallo-Hellenica 20 (1993): 191-197.
- Collingwood, C. A. "The Formicidae (Hymenoptera) of Fennoscandia and Denmark." Fauna Entomologica Scandinavica 8 (1979): 1-174.
- Collingwood, C.A. 1958. A survey of Irish Formicidae. Proceedings of the Royal Irish Academy 59B:213-219
- Comín del Río, P., and Andrés de Haro. "Datos iniciales para un estudio ecológico de las hormigas de Menorca." Bolletí de la Societat d´Historia Natural de les Balears 24 (1980): 23-48.
- Comín del Río, P., and X. Espadaler. "Ants of the Pityusic islands." In Biogeography of the Pityusic Islands, edited by H. Kuhbler, J. A. Alcover and C. Guerau, 278-231. The Hague: Junk, 1984.
- Comín del Río, P. "Los Formícidos de Menorca. Contribución al estudio taxonómico, geográfico y biológico." Tesina de licenciatura Universida (1977): 135 pp.
- Comín del Río, P. Estudio de los formícidos de Baleares: Contribución al estudio taxonómico, geográfìco y biológico. Palma de Mallorca: Universidad de las Islas Baleares, 1988.
- Consani M., and P. Zangheri. 1952. Fauna di Romagna. Imenotteri - Formicidi. Memorie della Societa Entomologica Italiana 31: 38-48.
- Csosz S., B. Marko, K. Kiss, A. Tartally, and L. Galle. 2002. The ant fauna of the Ferto-Hansag National Park (Hymenoptera: Formicidae). In: Mahunka, S. (Ed.): The fauna of the Fert?-Hanság National Park. Hungarian Natural History Museum, Budapest, pp. 617-629.
- Csősz S., B. Markó, and L. Gallé. 2001. Ants (Hymenoptera: Formicidae) of Stana Valley (Romania): Evaluation of the effectiveness of a myrmecological survey. Entomologica Romanica 6 : 121-126.
- Csősz S., B. Markó, and L. Gallé. 2011. The myrmecofauna (Hymenoptera: Formicidae) of Hungary: an updated checklist. North-Western Journal of Zoology 7: 55-62.
- Csősz S., H. C. Wagner, M. Bozsó, B. Seifert, W. Arthofer, B. C. Schlick-Steiner, F. Steiner, and Z Pénzes. 2014. Tetramorium indocile Santschi, 1927 stat. rev. is the proposed scientific name for Tetramorium sp. C sensu Schlick-Steiner et al. (2006) based on combined molecular and morphological evidence (Hymenoptera: Formicidae). Zoologischer Anzeiger 253: 469-481.
- Cuní, M. "Excursión entomológica y botánica a la Cerdaña española (Cataluña)." Anales de la Sociedad española de Historia Natural (1881): 377.
- Czechowski W., A. Radchenko, W. Czechowska and K. Vepsäläinen. 2012. The ants of Poland with reference to the myrmecofauna of Europe. Fauna Poloniae 4. Warsaw: Natura Optima Dux Foundation, 1-496 pp
- Czechowski, W., and A. G. Radchenko. "Formica lusatica Seifert, 1997 (Hymenoptera: Formicidae), an ant species new to Finland, with notes on its biology and the description of males." Myrmecologische Nachrichten 8 (2006): 257-262.
- De Stefani T. 1889. Miscellanea imenotterologica sicula. Nat. Sicil. 8: 140-145.
- Dekoninck W., F. Hendrickx, M. Dethier, and J. P. Maelfait. 2010. Forest Succession Endangers the Special Ant Fauna of Abandoned Quarries along the River Meuse (Wallonia, Belgium). Restoration Ecology 18(5): 681690.
- Della Santa E. 1995. Fourmis de Provence. Faune Provence 16: 5-37.
- Delye G. 1983. Contribution a l'etude des peuplements des invertebres de milieux extremes: les fourmis des dunes littorales del aCamargue (B. d. Rh) et de l'Espiguette (Gard). Bull. Soc. Linneenne Provence 35: 121-124.
- Dewes E. 2005. Ameisenerfassung im Waldschutzgebiet Steinbachtal/Netzbachtal. Abh. Delattinia 31: 89-118.
- Dlussky G. M., and B. Pisarski. 1970. Formicidae aus der Mongolei. Ergebnisse der Mongolisch-Deutschen Biologischen Expeditionen seit 1962, Nr. 46. Mitteilungen aus dem Zoologischen Museum in Berlin 46: 85-90.
- Donisthorpe H. 1914. Myrmecophilous notes for 1913. Entomologist's Record and Journal of Variation 26: 37-45.
- Donisthorpe, H. "A first instalment of the ants of Turkey." Annals and Magazine of Natural History (12)3 (1950): 1057-1067.
- Donisthorpe, H. "British ants, their life history and classification (2nd edn.)." London: G Routledge (1927): xvi + 436 pp.
- Du Merle P. 1978. Les peuplements de fourmis et les peuplements d'acridiens du Mont Ventoux II. - Les peuplements de fourmis. Terre Vie 32(1): 161-218.
- Dubovikoff D. A., and Z. M. Yusupov. 2018. Family Formicidae - Ants. In Belokobylskij S. A. and A. S. Lelej: Annotated catalogue of the Hymenoptera of Russia. Proceedingss of the Zoological Institute of the Russian Academy of Sciences 6: 197-210.
- Dusmet, J. M. "Linneo y los himenópteros de España." Linneo en España. Homenaje a su segundo centenario Zaragoza (1907): 475.
- Dvorak, L., P. BOGUSCH, I. MALENOVSKÝ, P. BEZDÌÈKA, K. BEZDÌÈKOVÁ, K. HOLÝ, P. LIKA, J. MACEK, L. ROLLER, M. RÍHA et al. "Hymenoptera of Hády Hill, near the city of Brno (Czech Republic), collected during the Third Czech-Slovak Hymenoptera meeting." Acta Musei Moraviae, Scientiae biologicae (Brno) 93 (2008): 53-92.
- Eidmann H. 1942. Zur Kenntnis der Ameisenfauna des Nanga Parbat. Zoologische Jahrbücher. Abteilung für Systematik, Ökologie und Geographie der Tiere 75: 239-266.
- El-Hawagry M. S., M. R. Sharaf, H. M. Al Dhafer, H. H. Fadl, and A. S. Aldawood. 2015. Addenda to the insect fauna of Al-Baha Province, Kingdom of Saudi Arabia with zoogeographical notes. Journal of Natural History http://dx.doi.org/10.1080/00222933.2015.1103913
- Else G., B. Bolton, and G. Broad. 2016. Checklist of British and Irish Hymenoptera - aculeates (Apoidea, Chrysidoidea and Vespoidea). Biodiversity Data Journal 4: e8050. doi: 10.3897/BDJ.4.e8050
- Emery C. 1897. Anhang. Verzeichniss der auf der zweiten Reise nach Kleinasien (1897) gesammelten Ameisen, mit einer Neubeschreibung. P. 239 in: Escherich, K. 1897. Zur Kenntniss der Myrmecophilen Kleinasiens. I. Coleoptera. Wiener Entomologische Zeitung 16: 229-239.
- Emery C. 1914. Wissenschaftliche Ergebnisse der Bearbeitung von O. Leonhard's Sammlungen. 5. Südeuropäische Ameisen (Hym.). Entomologische Mitteilungen. Berlin-Dahlem 3: 156-159.
- Emery C. 1916. Fauna entomologica italiana. I. Hymenoptera.-Formicidae. Bullettino della Società Entomologica Italiana 47: 79-275.
- Emery, C. "Alcune formiche dell'isola di Creta." Bull. Soc. Entomol. Ital. Resoc. Adun. 26 (1894): 7-10.
- Emery, C. "Alcune formiche di Malta." Bollettino della Società Entomologica Italiana 56 (1924): 11-12.
- Emery, C. "Beiträge zur Kenntniss der palaearktischen Ameisen." Öfversigt af Finska Vetenskaps-Societetens Förhandlingar (Helsinki) 20 (1898): 124-151.
- Emery, C. "Catalogo delle formiche esistenti nelle collezioni del Museo Civico di Genova. Parte seconda. Formiche dell'Europa e delle regioni limitrofe in Africa e in Asia." Annali del Museo Civico di Storia Naturale 12 (1878): 43-59.
- Emery, C. "Crociera del Violante, comandato dal capitano armatore Enrico d'Albertis, durante l'anno 1877. Formiche." Annali del Museo Civico di Storia Naturale 15 (1880): 389-398.
- Emery, C. "Voyage de M. Ch. Alluaud aux îles Canaries. Formicides." Annales de la Société Entomologique de France 62 (1893): 81-88.
- Entomological Society of Latvia. 2003. http://leb.daba.lv/Formicidae.htm (Accessed on December 1st 2013).
- Espadaler X. F. Garcia, K. Gomez, S. Serrano, and R. Vila. 2009. Ants (Hymenoptera, Formicidae) from Mont-Rebei gorge (Pallars Jussà). Boletín Sociedad Entomológica Aragonesa 44: 393399.
- Espadaler X., F. Garcia, X. Roig, and R. Vila. 2013. Ants (Hymenoptera, Formicidae) from the Castell de Montesquiu park (Osona, north-east of the Iberian Peninsula). Boletín de la Sociedad Entomológica Aragonesa (S.E.A.) 53: 223-227.
- Espadaler X., X. Roig, K. Gómez, and F. García. 2011. Formigues de les Planes de Son i mata de València (Hymenoptera, Formicidae) Treballs de la Institució Catalana d'Història Natural 16: 609-627.
- Espadaler, X. "Contribución al conocimiento de los formícidos (Hymenoptera, Formicidae) del Pirineo catalán." Tesis Universida (1979): 285 pp.
- Espadaler, X. "Formícidos de los alrededores de la laguna de Sariñena (Huesca). Descripción del macho de Camponotus foreli Emery." Colección Estudios Altoaragoneses 6 (1986): 109-126.
- Espadaler, X. "Lasius neglectus Van Loon, Boosma & Andrásfalvy, 1990 a potential pest ant in Spain." Orsis 14 (1999): 43-46.
- Espadaler. 2010. Hymenoptera (Formicidae) in Chapter 11: List of Arthropods (Arthropoda), in Borges P.A.V. and V Vieira et al. (Eds.). 2010. A list of the terrestrial and marine biota from the Azores. Principia, Oeiras, 432 pp.
- Eto S., and K. Ogata. 1983. Ants of Hirado Island, Kyushu. Bulletin of the Nagasaki Prefecture Biological Group 25: 7-11.
- Finzi, B. "Formiche dell'isola d'Elba e Monte Argentario." Bollettino della Società Entomologica Italiana 56 (1924): 12-15.
- Finzi, B. "Quarto contributo alla conoscenza della fauna mirmecologica della Venezia Giulia." Bollettino della Società Entomologica Italiana 60 (1928): 128-130.
- Finzi, B. "Risultati scientifici della spedizione Ravasini-Lona in Albania. III. Formiche." Bollettino della Società Entomologica Italiana 55 (1923): 1-4.
- Forel A. 1892. Die Ameisenfauna Bulgariens. (Nebst biologischen Beobachtungen.). 305-318.
- Forel A. 1893. Nouvelles fourmis d'Australie et des Canaries. Ann. Soc. Entomol. Belg. 37: 454-466.
- Forel A. 1900. Fourmis du Japon. Nids en toile. Strongylognathus Huberi et voisins. Fourmilière triple. Cyphomymrex Wheeleri. Fourmis importées. Mitt. Schweiz. Entomol. Ges. 10: 267-287.
- Forel A. 1904. Fourmis du Musée de Bruxelles. Ann. Soc. Entomol. Belg. 48: 168-177.
- Forel A. 1906. Fourmis d'Asie mineure et de la Dobrudscha récoltées par M. le Dr. Oscar Vogt et Mme Cécile Vogt, Dr. méd. Ann. Soc. Entomol. Belg. 50: 187-190.
- Forel A. 1907. Formiciden aus dem Naturhistorischen Museum in Hamburg. II. Teil. Neueingänge seit 1900. Mitt. Naturhist. Mus. Hambg. 24: 1-20.
- Forel A. 1911. Fourmis nouvelles ou intéressantes. Bull. Soc. Vaudoise Sci. Nat. 47: 331-400.
- Forel, A. "Ameisen aus den Sporaden, den Cykladen und Griechenland, gesammelt 1887 von Herrn von Oertzen." Berliner Entomologische Zeitschrift 32 (1889): 255-265.
- Forel, A. "Les Formicides de la Province d'Oran (Algérie)." Bulletin de la Société Vaudoise des Sciences Naturelles 30 (1894): 1-45.
- Forel, A. "Nouvelles fourmis d'Australie et des Canaries." Annales de la Société Entomologique de Belgique 37 (1893): 454-466.
- Forel, A. "Nouvelles fourmis de Grèce récoltées par M. E. von Oertzen." Annales de la Société Entomologique de Belgique 30 (1886): clix-clxviii.
- Formidabel Database
- Fowles, A.P. 1996. A provisional checklist of the invertebrates recorded from Wales. 2. Aculeate wasps, bees and ants (Hymenoptera: Aculeata). Countryside Council for Wales
- Franch, J., and X. Espadaler. "Ants as colonizing agents of pine stumps in San Juan de la Peña (Huesca, Spain)." Vie et Milieu 38 (1988): 149-154.
- Francois J. 1958. Contribution a l'etude ecologique des Formicides (Insectes, Hymenopteres) de la region Dijonnaise. Travaux du laboratoire de Zoologie et de la Station Aquicole Grimaldi de la Faculte des Sciences de Dijon 25, 39 pages.
- GRETIA. 2017. Bilan annuel de l'enquete sur la repartition des fourmis armoricaines. 23 pages.
- Gadeau de Kerville H. 1922. Materiaux pour la Faune des Hymenopteres de la Normandie. Bull. Soc. Amis Sc. Nat. Rouen 1916-1921, 1922: 217-225.
- Galle L. 1972. Study of ant-populations in various grassland ecosystems. Acta Biologica Szeged 18(1-4): 159-164.
- Galle L. 1981. The Formicoid fauna of the Hortobagy. Pp. 307-311 in: Mahunka, S. (ed.) 1981. The fauna of the Hortobágy National Park. Budapest: Akadémiai Kiadó, 415 pp.
- Galle L. 1997. Contribution to the ant fauna of Slovenia with special reference to the submediterranean and eudinaric regions. Annals for Istrian and Mediterranean studies 11: 209-214.
- Galle L., and G. Szonyi. 1988. A check list of ants (Hymenoptera: Formicoidea) of a sandy grassland in Kiskunsag National Park (Hungary). Acta Biol. Szeged 34: 167-168.
- Gallé L., B. Markó, K. Kiss, E. Kovács, H. Dürgő, K. Kőváry, and S. Csősz. 2005. Ant fauna of Tisza river basin (Hymenoptera: Formicidae). In: Gallé, L. (szerk.): Vegetation and Fauna of Tisza River Basin I. Tiscia Monograph Series 7; Szeged, pp. 149-197.
- Gaspar C. 1968. Les fourmis de la Drome et des Basses-Alpes, en France (Hymenoptera, Formicidae). Naturaliste can. 95: 747-766.
- Gaspar C., and C. Thirion. 1978. Modification des populations d'Hymenopteres sociaux dans les milieux anthropogenes. Memorabilia Zoologica 29: 61-77.
- Gaspare Charles. 1965. Étude myrmécologique d'une région naturelle de Belgique: la Famenne Survey des Fourmis de la Région (Hymenoptera, Formicidae). Institut agronomique de l'Etat a' Gembloux. 32(4): 427-434.
- Gasperini, R. "Notizia sulla fauna imenotterologadalmata, II, Formicidae-Mutillidae-Scoliadae-Sapygidae-Pompilidae-Sphegidae-Chrysididae." Anuario Dalmatico 4 (1887): 143-160.
- Ghahari H., C. A. Collingwood, M. Tabari, and H. Ostovan. 2009. Faunistic notes on Formicidae (Insecta: Hymenoptera) of rice fields and surrounding grasslands in northern Iran. Mun. Ent. Zool. 4(1): 184-189.
- Ghahari H., and C. A. Collingwood. 2013. A study on the ants (Hymenoptera: Vespoidea: Formicidae) from Western Iran. Acta Phytopathologica et Entomologica Hungarica 48 (1): 155164.
- Gholami M., A. Afshari, and S. A. Mafi Pashakolaei. 2012. The fauna and frequency of cottony cushion scale (Icerya purchasi Maskell) related ants (Hymenoptera: Formicidae) community in citrus orchards of Sari region, northern Iran. 20th Iranian Plant Protection Congress, 25-28 August 2012.
- Giacalone I., and M. Moretti. 2001. Contributo alla conoscenza della mirmecofauna (Hymenoptera: Formicidae) dei castagneti al Sud delle Alpi (ticino, Svizzera). Bollettino della Societa ticinese di Scienze naturali 89(1-2): 51-60.
- Gilev A. V., I. V. Kuzmin, V. A. Stolbov, and S. D. Sheikin. 2012. Materials on the fauna and ecology of ants (formicidae) Southern part of the Tyumen region. Tyumen State University Herald 6: 86-91.
- Gippet J. M., N. Mondy, J. Diallo-Dudek, A. Bellec, A. Dumet, L. Mistler, and B. Kauffmann. 2016. I’m not like everybody else: urbanization factors shaping spatial distribution of native and invasive ants are species-specific. Urban Ecosystems DOI 10.1007/s11252-016-0576-7
- Glaser F. 2009. Die Ameisen des Fürstentums Liechtenstein. (Hymenoptera, Formicidae). Amtlicher Lehrmittelverlag, Vaduz, 2009 (Naturkundliche Forschung im Fürstentum Liechtenstein; Bd. 26).
- Goetsch, W. "Beiträge zur Biologie spanischer Ameisen." EOS (Revista española de entomología) 18 (1942): 175-241.
- Gomez C., D. Casellas, J. Oliveras, and J. M. Bas. 2003. Structure of ground-foraging ant assemblages in relation to land-use change in the northwestern Mediterranean region. Biodiversity and Conservation 12: 21352146.
- Gomez K. 2017. Two species of exotic ants (Hymenoptera: Formicidae) new to Malta. Boletin de la Sociedad Entomologica Aragonesa (S.E.A.) 61: 233-235.
- González, A. L., J. A. Tinaut, and R. Gomez. "Evolución estacional de la entomofauna del suelo en un robledal (Quercus pyrenaica Wilde) de Sierra Nevada." Actas del III Congreso Ibérico de Entomología (1988): 557-576.
- Grandi G. 1935. Contributi alla conoscenza degli Imenotteri Aculeati. XV. Boll. R. Ist. Entomol. Univ. Studi Bologna 8: 27-121.
- Gratiashvili N., Barjadze S. 2008. Checklist of the ants (Formicidae Latreille, 1809) of Georgia. Proceedings of the Institute of Zoology (Tbilisi) 23: 130-146.
- Gribodo, G., Emery C. "Ordo Hymenoptera." Pp. 81-85 in: Cavanna, G. Parte II.- Catalogo degli animali raccolti al Vulture, al Pollino ed in altri luoghi dell'Italia meridionale e centrale Bull. Soc. Entomol. Ital. 14 (1882): 31-87.
- Groc S., J. H. C. Delabie, R. Cereghino, J. Orivel, F. Jaladeau, J. Grangier, C. S. F. Mariano, and A. Dejean. 2007. Ant species diversity in the Grands Causses (Aveyron, France): In search of sampling methods adapted to temperate climates. C. R. Biologies 330: 913922.
- Grzes I. M. 2009. Ant species richness and evenness increase along a metal pollution gradient in the Boles?aw zinc smelter area. Pedobiologia 53: 65-73.
- Guillen J. M. C., and A. Tinaut. 2012. Introduction to the study of the ants from the Region of Murcia (Spain) (Hymenoptera, Formicidae). Boln. Asoc. esp. Ent. 36 (1-2): 145-162.
- Gulzar A. 2014. Classification and Distribution of ants in Kashgar, Xinjiang. Master's Thesis Shaanxi Normal University, 75 pages.
- Guénard B., and R. R. Dunn. 2012. A checklist of the ants of China. Zootaxa 3558: 1-77.
- Gómez C., and S. Abril. 2011. Selective logging in public pine forests of the central Iberian Peninsula: Effects of the recovery process on ant assemblages Forest Ecology and Management 262: 1061-1066.
- Güsten, R., A. Schulz, and M. Sanetra. "Redescription of Tetramorium forte Forel, 1904 (Insecta: Hymenoptera: Formicidae), a western Mediterranean ant species." Zootaxa 1310 (2006): 1-35.
- Ha S.J, S.J. Park, and B.J. Kim. 2002. Comparative ant faunas between Seonyudo and seven other islands of West Sea in Korea. Korean Journal of Entomology 32(2): 75-79.
- Haguet G., M. Chevrier, and E. Brunel. 2002. Les invertebres de la dune de Bon Abri, premier inventaire. Groupe d'Etude des Invertebres Armoricains, 37 pages.
- Hayashida K. 1961. Studies on the ecological distribution of ants in Sapporo and its vicinity (1 et 2). Insectes Sociaux 7: 125-162.
- Hayashida K. 1964. Studies on the ecological distribution of ants in Kutchan and its adjacent area. Journal of the Sapporo Otani Junior College 2: 107-129.
- Hernandez Cuba O., J. L. Perez-Bote, and J. M. Garcia Jimenez. 2006. Los formicidos (Hymenoptera: Formicidae) del Parque Natural de Cornaldo (suroeste de la Peninsula Iberica). Boletin Sociedad Entomologica Aragonesa 38: 356-358.
- Hisamatsu M. 2004. List of Hymenoptera Recorded in Ibaraki Prefecture. Bulletin of Ibaraki Nature Museum 7: 125-164.
- Holgersen H. 1943. Ant studies in Rogaland (south-western Norway). Avhandlingar utgitt av det Norske Videnskaps-Akademi i Oslo. I. Matematisk-Naturvidenskapelig Klasse 1943(7): 1-75.
- Holgersen H. 1944. The ants of Norway (Hymenoptera, Formicidae). Nytt Magasin for Naturvidenskapene 84: 165-204.
- Hou J.H., D.W. Zhou, and S.C. Jiang. 2002. Species Composition and Spatial Distribution 0f Ants in the Grassland Region in the West of Jilin Province. Acta Ecologica Sinica 22(10): 1781-1787.
- IZIKO South Africa Museum Collection
- Ichikawa A. 1999. Records of ants observed from several localities of Osaka Prefecture, Japan, -1. Ari 23: 1-3.
- Ito. F., Kondoh. M., Kubota. S., Masuko. K., Morishita. M., Murata. K., Ogata. K., Sato. T., Takamine. H., Yamaoka. H. and Kondoh. M. 1986. A list of ants collected at Akiyoshi-dai (Yamaguchi-ken) by the members of the Myrmecologists Society (Japan) in 1985. ARI Reports of the Myrmecologists Society (Japan) 14: 5-6
- Izhaki I., B. Idelovich, R. Laster, and Y. Ofer. 2009. The impact of macro- vs micro environmental factors on the structure of ant communities inhabiting East-Mediterranean Aleppo pine forests. Israel Journal of Entomology 39: 129-146.
- Jeffery H. G. 1931. The Formicidae (or ants) of the Isle of Wight. Proceedings of the Isle of Wight Natural History and Archaeological Society 2: 125-128.
- Johnson R. Personnal Database. Accessed on February 5th 2014 at http://www.asu.edu/clas/sirgtools/resources.htm
- Kanizsai O., R. Gallé, and L. Gallé. 2009. Perception of spatial patchiness by ant assemblages (Hymenoptera: Formicidae). Tiscia 37: 3-7.
- Karaman M. G. 2009. An introduction to the ant fauna of Macedonia (Balkan Peninsula), a check list (Hymenoptera, Formicidae). Natura Montenegrina 8(3): 151-162.
- Karaman M. G. 2011. A catalogue of the ants (Hymenoptera, Formicidae) of Montenegro. Podgorica: Catalogues 3, Volume 2, Montenegrin Academy of Sciences and Arts, 140 pp.
- Karaman M. G., and G. S. Karaman. 2003. Contribution to the knowledge of the ants (Hymenoptera, Formicidae) from Serbia. The Montenegrin Academy of Sciences and Arts. Glasnik of the Section of Natural Sciences 15: 39-58.
- Karaman M. G., and G. S. Karaman. 2005. New data on myrmecofauna (Hymenoptera, Formicidae) from Montenegro (Serbia & Montenegro). The Montenegrin Academy of Sciences and Arts. Glasnik of the Section of Natural Sciences 16: 51-68.
- Karavaiev V. 1910. Nachtrag zu meinen Ameisen aus Transcaspien und Turkestan. Rus. Entomol. Obozr. 9: 268-272.
- Karavaiev V. 1911. Ameisen aus Transkaspien und Turkestan. Tr. Rus. Entomol. Obshch. 39: 1-72.
- Karavaiev V. 1912. Ameisen aus dem paläarktischen Faunengebiete. Rus. Entomol. Obozr. 12: 581-596.
- Karavaiev V. 1926. Beiträge zur Ameisenfauna des Kaukasus, nebst einigen Bemerkungen über andere palaearktische Formen. (Fortsetzung). Konowia 5: 161-169.
- Karavaiev V. 1927. Übersicht der Ameisenfauna der Krim nebst einigen Neubeschreibungen. Konowia 5: 281-303.
- Kawahara Y., S. Hosoyamada, and S. Yamane. 1999. Ant fauna of the Terayama Station for Education and Research on Nature, Kagoshima University. Bulletin of the Faculty of Education, Kagoshima University. Natural Science 50: 147-156.
- Kempf, W.W. 1972. Catalago abreviado das formigas da regiao Neotropical (Hym. Formicidae) Studia Entomologica 15(1-4).
- Khamraev A. S. 2003. Soil organisms and entomocomplexes in Khorezm and Karakalpakstan (Uzbekistan). ZEF work paper for sustainable development in Central Asia No 6, 71 pages
- Kim B., Ryu D., Park S., and J. Kim. 1994. Systematic study on ants from coasts of Korean Peninsula (Hym: Formicidae). Korean journal of entomology 24: 293-309.
- Kim B., S. Park, J. Kim, K. Kim. 1997. Ants from Mount Moak (Hymenoptera: Formicidae). Korean Journal of Soil Zoology 2(1): 41-58.
- Kim B.J. 1986. A systematic study of ants in Island Ullungdo of Korea on the basis of external fine features. The journal of Natural Science 5(2): 84-94.
- Kim B.J. 1996. Synonymic list and distribution of Formicidae (Hymenoptera) in Korea. Entomological Research Bulletin Supplement 169-196.
- Kim B.J., K.G. Kim, D.P. Ryu, J.H. Kim. 1995. Ants of Chindo island in Korea (Hymenoptera; Formicidae). The Korean Journal of Systematic Zoology 11(1): 101-113.
- Kim B.J., K.G. Kim, J.H. Kim, S.J. Park. Ants from Mt. Mirok. Korean J. Soil Zoology 2(2): 115-128.
- Kim B.J., S.J. Park, J.H. Kim, K.G. Kim. 1997. Ants from Mt. Moak (Hym., Formicidae) Korean J. Soil Zoology 2(1): 41-58.
- Kim B.J., S.J. Park, and J.H. Kim. 1996. Ants from Naejangsan national park (Hymenoptera: Formicidae). Korean J. Soil. Zoology &(2): 120-133.
- Kim B.J.; Kim, K.G.; Lim, K.H. 1993. Systematic study of ants from Chejudo province. Korean Journal of Entomology 23: 117-85
- Kim B.J.; Kim, K.G.; Lim, K.H. 1993. Systematic study of ants from Chejudo province. Korean Journal of Entomology 23: 117-86
- Kim B.J.; Kim, K.G.; Lim, K.H. 1993. Systematic study of ants from Chejudo province. Korean Journal of Entomology 23: 117-87
- Kim B.J.; Kim, K.G.; Lim, K.H. 1993. Systematic study of ants from Chejudo province. Korean Journal of Entomology 23: 117-88
- Kim B.J.; Kim, K.G.; Lim, K.H. 1993. Systematic study of ants from Chejudo province. Korean Journal of Entomology 23: 117-89
- Kim B.J.; Kim, K.G.; Lim, K.H. 1993. Systematic study of ants from Chejudo province. Korean Journal of Entomology 23: 117-90
- Kim B.J.; Kim, K.G.; Lim, K.H. 1993. Systematic study of ants from Chejudo province. Korean Journal of Entomology 23: 117-91
- Kim B.J.; Kim, K.G.; Lim, K.H. 1993. Systematic study of ants from Chejudo province. Korean Journal of Entomology 23: 117-92
- Kim B.J.; Kim, K.G.; Lim, K.H. 1993. Systematic study of ants from Chejudo province. Korean Journal of Entomology 23: 117-93
- Kim B.J.; Kim, K.G.; Lim, K.H. 1993. Systematic study of ants from Chejudo province. Korean Journal of Entomology 23: 117-94
- Kim B.J.; Kim, K.G.; Lim, K.H. 1993. Systematic study of ants from Chejudo province. Korean Journal of Entomology 23: 117-95
- Kim C.H., B.M. Choi, and J.R. Bang. 1992. Studies on the distribution of ants (Formicidae) in Korea (8)-Ant fauna in 10 islands, Chollanam-do. Korean J. Appl. Entomol. 31(4): 345-359.
- Kim C.H.; Choi, B.M. 1987. On the kinds of ants (Hymenoptera: Formicidae) and vertical distribution in Jiri Mountain. Korean Journal of Plant Protection 26:123-132.
- Kim K.I., C.H. Kim, and B. Choi. 1989. The ant fauna of the southern shore in Gyeongsangnamdo, Korea. Journal of Gyeongsang Nat. Univ. 28(2): 213-226.
- Kim et al. 1993. Systematic study of ants from Chejudo Province. Koran Journal of Entomology 23(3): 117-141.
- Kim, Byung-Jin, Ky-Gyong Kim, Dong-Pyo Ryu and Joong-Hyon Kim. 1995. The Korean Journal of Systematic Zoology. 11(1):101-113.
- Kiran K., C. Karaman, and V. Aksoy. 2014. Strongylognathus (Hymenoptera, Formicidae) cinsi kar?ncalarda atipik sosyal parazitlik. Proceeding of 22st National Congress of Biology, Eski?ehir, Turkey.
- Kiran K., and C. Karaman. 2012. First annotated checklist of the ant fauna of Turkey (Hymenoptera: Formicidae). Zootaxa 3548: 1-38.
- Kiran K., and N. Aktac. 2006. The vertical distribution of the ant fauna (Hymenoptera: Formicidae) of the Samanh Mountains, Turkey. Linzer Biol. Beitr. 38(2): 1105-1122.
- Kiss K., and K. Fetyko. 2008. Notes about the ant fauna (Hymenoptera: Formicidae) of the Tur valley. In. Sike T., Márk-Nagy J. (eds), The flora and fauna of the Tur river natural reserve, Biharean Biologist, II, Oradea, pp.71-76.
- Kofler A. 1995. Nachtrag zur Ameisenfauna Osttirols (Tirol, Österreich) (Hymenoptera: Formicidae). Myrmecologische Nachrichten 1: 14-25.
- Kondoh M., and Y. Kitazawa. 1984. Ant communities of the campus of UOEH and in an adjacent natural forest. Journal of UOEH 6(3): 221-234.
- Korlevic, A. "Prilozi fauni hrvatskih opnokrilaca." Glasn. Hrv. Narav. Dr. 5 (1890): 189-250.
- Kozisek T. 1987. Ants (Formicoidea) of the Abrod State Nature Reserve. Ochrana Prírody 8: 205-208.
- Kratochvíl J., V. Novák, and J. Snoflák. 1944. Mohelno. Soubor práci venoványch studiu vyznamne památky prírodní. 5. Hymenoptera - Aculeata. Formicidae - Apidae - Vespidae. Arch. Svazu Ochr. Prír. Domov. Moravé 6: 1-155.
- Krausse, A. H. "Ueber sardische Ameisen." Archiv für Naturgeschichte, Berlin A 78(7) (1912): 162-166.
- Kubota M. 1983. Records of ants (3). Ari 11: 7-8.
- Kubota S., and M. Terayama. 1982. Ant fauna of Kanagawa Prefecture, Japan (IV) Ants of Kakio. Kanagawa-chuho (Journal of the Kanagawa Entomologists Association) 21-28.
- Kubota S.; Terayama, M. 1989. Ant fauna of Tokyo. (1) A list of ants collected at the parks. Ari 16:14-16.
- Kubota. S., and M. Terayama. 1988. Ant fauna of Tokyo. (1) A list of ants collected at the parks. ARI Reports of the Myrmecologists Society (Japan) 16: 14-16
- Kugler J. 1988. The zoogeography of Israel. 9. The zoogeography of social insects of Israel and Sinai. Monographiae biologicae 62: 251-275.
- Kvamme T. 1982. Atlas of the Formicidae of Norway (Hymenoptera: Aculeata). Insecta Norvegiae 2: 1-56.
- Kvamme T., and A. Wetas. 2010. Revidert liste over norske maur Inkludert dialektiske navn og forslag til nye norske navn og forslag til norske navn. Norsk institutt for skog og landskap, Ås. 127 pp
- Kwon T. S. 2012. Korean ant atlas. Korea Forest Research Institute 162 pages.
- Kwon T. S. 2015. Ant assemblages along the Baekdudaegan Mountain Range in South Korea: Human roads and temperature niche. Journal of Asia-Pacific Biodiversity 8: 152-157.
- Kwon T.-S., M.-Y. Song, S.-C. Shin, and Y.-S. Park. 2005. Effects of aerial insecticide sprays on ant communities to control pine wilt disease in Korean pine forests. Appl. Entomol. Zool. 40(4): 563574.
- Laeger T., and R. Schultz. 2005. Ameisen (Hymenoptera: Formicidae) als Beifänge in Bodenfallen wie genau spiegeln sie reale Abundanzverhältnisse wider? Myrmecologische Nachrichten 7: 17-24.
- Legakis A. 1983. Aspects of the distribution of ants in an insular mediterranean ecosystem (Cyclades Is., Greece) Rapports et Procès-Verbaux des Réunions de la Commission Internationale pour l'Exploration Scientifique de la Mer Méditerranée. Monaco 28: 121-122.
- Legakis A. 1983. The Zoological Museum of the University of Athens 2. The collection of ants from Greece. Biologia Gallo-Hellenica 11(1): 85-87.
- Legakis A. 1989. Aspects of the distribution of ants in an insular mediterranean ecosystem (Cyclades Isl., Greece). Rev. Ecol. Biol. Sol. 26 (3): 363-369.
- Legakis A. 2001. Ants (Hymenoptera, Formicidae) collected in the eastern Pyrenees and surrounding area, September 1999. Proceedings of the International Colloqium of the European Invertebrate Survey: Cartography and Conservation of Biodiversity Marcevol Priory, Arboussols, france 30.8-4.9.1999. OPIE.
- Legakis Collection Database
- Lehouck V., D. Bonte, W. Dekoninck, and J. P. Maelfait. 2004. The distribution of ant nests (Hymenoptera, Formicidae) in coastal grey dunes of Flanders (Belgium) and their relationship to myrmecochorous plants. Belg. J. Zool. 134 (2/1) : 89-96.
- Lenoir A. 1971. Les fourmis de Touraine, leur intérêt biogéographique. Cahiers des Naturalistes 27: 21-30.
- Li Z.h. 2006. List of Chinese Insects. Volume 4. Sun Yat-sen University Press
- Liu H., X.Z. Yuan, C.D. Zhang. 2002. Community structure and species diversity of ant in Qufu, Shandong province. Biodiversity Science 10(3): 298-304.
- Livory A. 2003. Les fourmis de la Manche. L'Argiope 39: 25-49.
- Livory A. Un Nouvel Inventaire: les Fourmis. Argiope 29: 27-34.
- Lomnicki J. 1928. Spis mrówek Lwowa i okolicy. Ksiegi Pamiatkowej (Lecia Gimn. IV Jana Dlugosza Lwowie) 50: 1-10.
- Lomnicki, J. "Une contribution à la connaissance de la faune des fourmis des îles Baléares." Polskie Pismo Entomologiczne - Bulletin Entomologique de Pologne 4 (1925): 1-3.
- Lopez, F., J. M. Zorrilla, F. J. Acosta Salmerón, and J. M. Serrano. "Comparative morphological study of Tetramorium caespitum (Linné, 1758) and Tetramorium semilaeve André, 1881 (Hym., Formicidae)." Miscellania Zoologica (Barcelona) 15 (1992): 169-178.
- Lorite P., J. A. Carrillo, M. F. García, and T. Palomeque. 2000. Chromosome numbers in Spanish Formicidae. III. Subfamily Myrmicinae (Hymenoptera). Sociobiology 36: 555-570.
- López, F., J. M. Serrano, and F. J. Acosta. "Compared Iberian distribution of Tetramorium caespitum (Linné, 1758) and Tetramorium semilaeve André, 1881 (Hym., Formicidae)." Anales de Biología 16 (1990): 53-61.
- López, F. "Estudio morfológico y taxonómico de los grupos de especies ibéricas del género Tetramorium Mayr, 1855." Boletín de la Asociación Española de Entomología 15 (1991): 29-52.
- López, F. "Variabilidad morfológica y problemas taxonómicos en Tetramorium caespitum (Linné 1758) y Tetramorium semilaeve André, 1881." Boletín de la Asociación Española de Entomología 15 (1991): 65-78.
- Maavara V. 1953. Ants of Estonian SSR. ABIKS loodusevaatlejale 10: 1-44.
- Manabe K. 1994. Ants of the shrine forest in Fukuoka Prefecture (First report; Ants of lowland). Ari 17: 6.
- Manabe K. 1994. Ants of the shrine forest in Fukuoka Prefecture (first report; ants of lowland). Ari 18: 6.
- Marko B. 1999. Contribution to the knowledge of the myrmecofauna of the River Somes valley. In: Sárkány-Kiss, A., Hamar, J. (szerk.): The Somes/Szamos River Valley. A study of the geography, hydrobiology and ecology of the river system and its environment. Tiscia Monograph Series 3, Szolnok-Szeged-Tîrgu Mure?, pp. 297-302.
- Markó B. 1997. Contribution to the Knowledge of the Ant-Fauna (Hymenoptera: Fromicoidea) of the Cri?ul-Repede River-Valley. In: Sárkány-Kiss, A., Hamar, J. (szerk.): The Cri?/Körös Rivers' Valleys. A study of the geography, hydrobiology and ecology of the river system and its environment. Tiscia Monograph Series 2, Szolnok-Szeged-Tîrgu Mure?, pp. 345-352.
- Markó B., and S. Csősz. 2002. Die europäischen Ameisenarten (Hymenoptera: Formicidae) des Hermannstädter (Sibiu, Rumänien) Naturkundemuseums I.: Unterfamilien Ponerinae, Myrmicinae und Dolichoderinae. Annales Historico-Naturales Musei Nationalis Hungarici 94: 109-121.
- Martorell, M. "Catálogos sinonímicos de insectos encontrados en Cataluña." Barcelona (1879).
- Martínez Ibáñez, M. D., and A. Tinaut. "Nuevas especies de formícidos (Hymenoptera, Formicidae) para la Sierra de Albarracín (Teruel)." Real Sociedad Española de Historia Natural Tomo extra (1996): 174-177.
- Martínez Ibáñez, M. D., and J. M. Serrano Talavera. "Contribución al conocimiento de las hormigas (Hym. Formicidae) del sabinar español." Boletim da Sociedade Portuguesa de Entomologia Suplemento Actas Do I (1985): 33-41.
- Masuko K. 1986. A list of ants collected at Akiyoshi-dai (Yamaguchi-ken) by the members of the Myrmecologists Society (Japan) in 1985. Ari 14:5.
- Medina, M. "Catálogo provisional de las hormigas de Andalucía." Actas de la Sociedad Española de Historia Natural 20 (1891): 95-104.
- Mei M. 1995. Arthropoda di Lampedusa, Linosa e Pantelleria (Canale di Sicilia, Mar Mediterraneo). Hymenoptera Formicidae (con diagnosi di due nuove specie). Il Naturalista Siciliano (4)19(suppl.): 753-772.
- Menozzi C. 1918. Primo contributo alla conoscenza della fauna mirmecologica del Modenese. Atti della Società dei Naturalisti e Matematici di Modena. (5)4: 81-88.
- Meyer M., C. Braunert, R. Gerend, and I. Schrankel. 2003. Résultats de l'excursion annuelle du groupe de travail entomologique de la Société des naturalistes luxembourgeois. Bull. Soc. Nat. luxemb. 103: 103-120.
- Minato M., T. Kameyama, F. Ito, and T. Itino. 1996. A preliminary report of ant fauna in Gagawa Prefecture. Ari 20: 9-13.
- Mizota K. 2002. A check list of insects in Kinkazan Island, Miyagi Pref., Northeastern Japan: A bibliographical Survey. Bulletin of Miyagi University of Education Environmental Education 5: 69-78.
- Moretti M. P. Duelli, and M. K. Obrist. 2006. Biodiversity and resilience of arthropod communities after fire disturbance in temperate forests. Oecologia 149: 312327.
- Morillo C. 1981. Inventario y fenologia de los Formicidae de un encinar de Espana central. Rev. Ecol. Biol. Sol. 18(2): 243-251.
- Müller, G. "Le formiche della Venezia Guilia e della Dalmazia." Bollettino della Società Adriatica di Scienze Naturali in Trieste 28 (1923): 11-180.
- Nadig A. 1918. Alcune note sulla fauna dell'alta Valsesia. Formicidae. Atti Soc. Ital. Sci. Nat. Mus. Civ. Stor. Nat. Milano 56: 331-341.
- Natuhara Y. 1998. Ant faunae in Osaka City and three other sites in Osake Prefecture. Bulletin of Myrmecological Society of Japan 22: 1-5.
- Negoro H. 1994. Ants from Toyama Prefecture, Hokuriku. Bulletin of the Toyama Science Museum 17: 35-47.
- Neumeyer R., and B. Seifert. 2005. Commented check list of free living ant (Hymenoptera: Formicidae) species of Switzerland. Bulletin de la Societe Entomologique Suisse 78: 1-17.
- Nielsen M. G. 2011. A check list of Danish ants and proposed common names. Ent. Meddr. 79: 13-18.
- Nielsen, M.G. 1986. Respiratory rates of ants from different climatic areas. Journal of Insect Physiology 32(2): 125-131
- Nieves-Aldrey J. L., F. Fontal-Cazalla, A. M. Garrido-Torres, and C. Rey del Castillo. 2003. Inventorio de Hymenoptera (Hexapoda) en el ventorrillo: un rico enclave de biodiversidad en la Sierra de Guadarrama (Espana Central). Graellsia 59(2-3): 25-43.
- Noordijk, J., R. Morssinkhof, P. Boer, A. P. Schaffers, Th. Heijerman and K. V. Sýkora. 2008. How ants find each other; temporal and spatial patterns in nuptial flights. Insectes Sociaux 55(3):266-273.
- Nylander, W. "Synopsis des Formicides de France et d'Algérie." Annales des Sciences Naturelles, Zoologie (4)5 (1856): 51-109.
- O'Rourke F. J. 1948. The distribution and general ecology of the Irish Formicidae. Proceedings of the Royal Irish Academy. Section B: Biological, Geological, and Chemical Science 52: 383-410.
- Ochi K. 1983. Distribution pattern of ants in pine stands, with special reference to Monomorium nipponense Wheeler (Hymenoptera: Formicidae). Gensei 44:1-6.
- Odegaard F., K. M. Olsen, A. Staverlokk, and J. O. Gjershaug. 2015. Towards a new era for the knowledge of ants (Hymenoptera, Formicidae) in Norway? Nine species new to the country. Nowegian Journal of Entomology 62: 80–99.
- Ogata K. 1981. The ant fauna of the Goto islands, Natural history of the Goto? Islands, Japan : Iki Tsushima to no taihi (Danjo Gunto? Ko?rai Sone o fukumu Japan: 347-351.
- Ogata K., Y. Hirashima, T. Miura, Y. Maeta, K. Yano, and J. Ko. 1985. Ants collected in pine forests infested by the pine needle gall midge in Korea (Hymenoptera, Formicidae). Esakia (23): 159-163.
- Ogata. K., Touyama, Y. and Choi, B. M. 1994. Ant fauna of Hiroshima Prefecture, Japan. ARI Reports of the Myrmecologists Society (Japan) 18: 18-25
- Ohkawara K., and Y. Fukushima. 1998. A Preliminary report of ant fauna in Ishikawa Prefecture, Central Japan. Ari 22: 6-9.
- Omid P., and H. G. Kami. 2007. New and additional records for the formicid fauna (Insecta: Hymenoptera) of Iran. Zoology in the Middle East 40(1): 85-90.
- Ortiz, F. J. "Formícidos del litoral granadino." Memoria de Licenciatura Universida (1985): 206 pp.
- Ottonetti L., L. Tucci, and G. Santini. 2006. Recolonization Patterns of Ants in a Rehabilitated Lignite Mine in Central Italy: Potential for the Use of Mediterranean Ants as Indicators of Restoration Processes. Restoration Ecology 14(1): 6066.
- Ovazza M. 1950. Contribution à la connaissance des fourmis des Pyrénées-Orientales. Récoltes de J. Hamon Vie et Milieu 1: 93-94.
- Ovazza M. 1953. Contributions a l'etude biologique de la Camargue , Formicides. Vie et Milieu 4(4) : 751-753.
- Palanca, A., A. Urbieta, C. Castán, and X. Espadaler. "Ants in the supraforestal pastures of the Tendeñera massif." Écologie des milieux montagnards et de haute altitude. Documents d'Ecologie Pyrénéenne III-IV (1984): 223-224.
- Paraschivescu D. 1972. Fauna mirmecologica din zonele saline ale Romaniei. Studii si Cercetari de Biologie. Seria Zoologie 24: 489-495.
- Paraschivescu D. 1978. Elemente balcanice in mirmecofauna R. S. Romania. Nymphaea 6: 463- 474.
- Park S.J., K.G. Kim, J.H. Kim, B.J. Kim. 1998. Ants from Chiaksan National Park. Korean J. Soil Zoology 3(1): 25-34.
- Park S.J., K.G. Kim, J.H. Kim, and B.J. Kim. 1998. Ants from the Seoraksan National Park. The Korean Journal of Systematic Zoology 14(4): 429-439.
- Park S.J., and B.J. Kim. 2002. Faunal comparison of ants among Cheongsando and other islands of South Sea in Korea. Korean Journal of Entomology 32(1): 7-12.
- Park, Seong, Joon and Byung, and Kim, Jin. 2002. Faunal Comparison of Ants among Cheongsando and Other Islands of South Sea in Korea. Korean Jornal of Entomology. 32(1):7-12.
- Pascual, M. R. "Estudio taxonómico y ecológico de los Formícidos de las Sierras de Alfacar, La Yedra, Huétor y Harana." Tesis Doctoral Universida (1986): 264 pp.
- Pashaei Rad S., B. Taylor, R. Torabi, E. Aram, G. Abolfathi, R. Afshari, F. Borjali, M. Ghatei, F. Hediary, F. Jazini, V. Heidary Kiah, Z. Mahmoudi, F. Safariyan, and M. Seiri. 2018. Further records of ants (Hymenoptera: Formicidae) from Iran. Zoology in the Middle East 64(2): 145-159.
- Passera L. 1967. Peuplement en fourmis terricoles du rebord Meridional des Causses jurassiques du Quercy: la lande calcaire a buis. Vie et milieu. Serie C, Biologie terrestre 18(1): 189-205.
- Passera L. 1977. Peuplement myrmecologique du cordon littoral du Languedoc-Roussillon modifications anthropiques. Vie et Milieu 27: 249-265.
- Pavlova N. S. 2014. To ant fauna (Hymenoptera, Formicidae) of the floodplain of Medvedista River (Saratov Province). Entomological and parasitological studies in the Volga region 11: 145-147.
- Pekar, S. 2004. Predatory Behavior of Two European Ant-Eating Spiders (Araneae, Zodariidae). Journal of Arachnology 32(1): 31-41
- Peng Y. 2008. Ants resources in Heilongjiang Province and Flora (Hymenoptera). Master's Thesis Northeast Normal University, China. 90 Pages.
- Petal J. 1974. Analysis of a sheep pasture ecosystem in the Pieniny mountains (the Carpathians) XV. The effect of pasture management on ant population. Ekologia Polska 22(3/4): 679-692.
- Petal J. M. 1967. Contribution a la connaissance des fourmis (Formicidae, Hymenoptera) de la region de Lublin. VII. Associations des fourmis des milieux de tourbieres, de forets et de dunes aux environs de Libiszow (dist. De Parczew). Annales Universitatis Mariae Curie-Sklodowska Lublin-Polonia 22(9): 117-130.
- Petrov I. Z. 2002. Contribution to the myrmecofauna (Formicidae, Hymenoptera) of the Banat Province (Serbia). Archives of Biological Sciences, Belgrade, 54(12): 57-64.
- Petrov I. Z. 2010. Contribution to the myrmecofauna (Hymenoptera: Formicidae) of Krusevac and its surroundings. Acta entomologica serbica, 2010, 15(1): 145-148.
- Petrov I. Z. 2012. Preliminary data on ants (Formicidae, Hymenoptera) on Mountain Avala (Belgrade, Serbia). Bulletin of the Natural History Museum 5: 95-99.
- Petrov I. Z., B. Petrov, D. Milicic, T. Karan-Znidarsic. 2007. Contribution to the Myrmecofauna (Hymenoptera: Formicidae) of East and South Serbia. Acta Zoologica Bulgarica 59(3): 295-299.
- Petrov I. Z., and C. A. Collingwood. 1992. Survey of the myrmecofauna (Formicidae, Hymenoptera) of Yugoslavia. Archives of Biological Sciences (Belgrade) 44: 79-91.
- Pisarski B. 1966. Études sur les fourmis du genre Strongylognathus Mayr. Annales Zoologici, Warszawa. 23: 509-523.
- Plateaux L. 2000. Les fourmis de Lorraine. Bull. Soc. Ent. Lorr. 11-16.
- Poldi B., M. Mei, and F. Rigato. 1995. Hymenoptera, Formicidae. Vol 102. Checklist delle specie della fauna Italiana: 1-10.
- Pusvaskyte O. 1979. Myrmecofauna of the Lituanian SSR. Acta Entomologica Lituanica 4: 99-105.
- Radchenko A. G. 2007. The ants (Hymenoptera, Formicidae) in the collection of William Nylander. Fragmenta Faunistica (Warsaw) 50: 27-41.
- Retana J., and X. Cerdá X. 2000. Patterns of diversity and composition of Mediterranean ground ant communites tracking spatial and temporal variability in the thermal environment. Oecologia 123: 436-444.
- Reyes-López J., S. Carpintero-Ortega, and E. Retamoza-Muñoz. 2010. Adiciones a la relación de especies de hormigas (Hym., Formicidae) del Parque Natural del Cabo de Gata- Níjar (Almería, España) Boletín de la Asociación Española de Entomología 34: 67-76.
- Reznikova Z. I. 2003. Distribution patterns of ants in different natural zones and landscapes in Kazakhstan and West Siberia along a meridian trend. Euroasian Entomological Journal 2(4): 235-342.
- Rigato F., and J. K. Wetterer. 2018. Ants (Hymenoptera: Formicidae) of San Marino. Natural History Sciences. Atti Soc. it. Sci. nat. Museo civ. Stor. nat. Milano, 5(2): https://doi.org/10.4081/nhs.2018.367
- Rigato F., and R. Sciaky. 1989. Contributo alla conoscenza della mirmecofauna della Val Gesso (alpi Marittime) (Hymenoptera, Formicidae). Boll. Mus. Reg. Sci. Nat. Torino 7(2): 427-442.
- Rigato F., and R. Sciaky. 1991. The myrmecofauna of the Gesso Valley (Maritime Alps) (Hymenoptera Formicidae). Ethology Ecology and Evolution Special Issue 1: 87-89.
- Rigato S., and I. Toni. 2011. Short notes 21. Hymenoptera, Formicidae. Pp. 873-882 in: Nardi, G.; Whitmore, D.; Bardiani, M.; Birtele, D.; Mason, F.; Spada, L.; Cerretti, P. (eds.) 2011. Biodiversity of Marganai and Montimannu (Sardinia). Research in the framework of the ICP Forests network. Conservazione Habitat Invertebrati, 5. Sommacampagna, Verona: Cierre Edizioni, 896 pp.
- Rodríguez, A. "Contribución al conocimiento de las hormigas (Hymenoptera, Formicidae) de Sierra Morena Central." Boletín de la Asociación Española de Entomología 5 (1982): 181-188.
- Rosenhauer, W. G. "Die Thiere Andalusiens." Erlangen (1856).
- Ruzsky M. 1902. Contribution to the ant fauna of the Turgai region. Rus. Entomol. Obozr. 2: 232-235.
- Ruzsky M. 1916. On zoological research in Yeniseisk province, work of summer of 1915. Izv. Imp. Tomsk. Univ. 65 (3rd p part: 1-21.
- Ruzsky M. 1946. Ants of Tomsk province and contiguous localities. Tr. Tomsk. Gos. Univ. 97: 69-121
- Röszler P. 1950. Die Ameisenwelt des Nagy Pietrosz, 2305 m (Ungarn) und Umgebung. Zool. Anz. 145: 210-225.
- Saaristo M. I. 1995. Distribution maps of the outdoor myrmicid ants (Hymenoptera, Formicidae) of Finland, with notes on their taxonomy and ecology. Entomol. Fennica 6: 153-162.
- Salata S. 2014. Ants (Hymenoptera: Formicidae) of the National Park of the Stołowe Mts. Przyroda Sudetow 17: 161-172.
- Salata S., and L. Borowiec. 2018. A new species of the ant genus Lasius Fabricius, 1804 from Crete (Hymenoptera, Formicidae). ZooKeys 789: 139–159.
- Salata S., and L. Borowiec. 2019. Comments to distribution of several Greek Tetramorium Mayr, 1855 species (Hymenoptera: Formicidae). Annals of the Upper Silesian Museum in Bytom Entomology 28(2): 1-9.
- Salgueiro, J. "Variaçao anual em três comunidades de formicídeos da Serra da Estrela. Adiçao de um género novo e de duas espécies novas para portugal." Boletín de la Asociación Española de Entomología 26 (3-4) (2002): 121-131.
- Sandor C., and M. Balint. 2004. Redescription of Tetramorium hungaricum RÖSZLER, 1935, a related species of T. caespitum (Linnaeus , 1758) (Hymenoptera: Formicidae). Myrmecologische Nachrichten 6: 49-59.
- Sanetra M., R. Güsten, and A. Schulz. 1999. On the taxonomy and distribution of Italian Tetramorium species and their social parasites (Hymenoptera Formicidae). Memorie della Societa Entomologica Italiana 77: 317-357.
- Santschi F. 1910. Contributions à la faune entomologique de la Roumanie. Formicides capturées par Mr. A. L. Montandon. Buletinul Societatii Române de Stiinte 19: 648-652.
- Schar S., G Talavera, X. Espadaler, J. D. Rana, A. A. Andersen, S. P. Cover, and R. Vila. 2018. Do Holarctic ant species exist? Trans-Beringian dispersal and homoplasy in the Formicidae. Journal of Biogeography 00: 1-12.
- Schembri, Stephen P. and Cedric A. Collingwood. A Revision of the Myrmecofauna of the Maltese Islands. 417-442.
- Schifani E., and A. Alicata. 2018. Exploring the myrmecofauna of Sicily: thirty-two new ant species recorded, including six new to Italy and many new aliens (Hymenoptera, Formicidae). Polish Journal of Entomology 87 (4): 323–348.
- Schlick-Steiner B. C., and F. M. Steiner. 1999. Faunistisch-ökologische Untersuchungen an den freilebenden Ameisen (Hymenoptera: Formicidae) Wiens. Myrmecologische Nachrichten 3: 9-53.
- Schmitz, H. "Ein Verzeichnis portugiesischer Ameisen (Formicidae, Hymenoptera)." Brotéria Ser. Cienc. Nat. 24 (1955): 27-37.
- Schulz, A., and M. Sanetra. "Notes on the socially parasitic ants of Turkey and the synonymy of Epimyrma (Hymenoptera: Formcidae)." Entomofauna, Zeitschrift für Entomologie 23 (14) (2002): 157-172.
- Scupola A., and R. Poggi. 2014. Ricerche zoologiche della nave oceanografica "minerva" (C.N.R.) sulle isole circumsarde. XXXIII. Formiche. Estratto dagli Annali del Museo Civico di Storia Naturale “G. Doria” 106: 93-131.
- Seifert B. 1994. Die freilebenden Ameisenarten Deutschlands (Hymenoptera: Formicidae) und Angaben zu deren Taxonomie und Verbreitung. Abhandlungen und Berichte des Naturkundemuseums Görlitz 67(3): 1-44.
- Shiran E., M. S. Mossadegh, and M. Esfandiari. 2012. Mutualistic ants (Hymenoptera: Formicidae) associated with aphids in central and southwestern parts of Iran. J. Crop Prot. 2013, 2 (1): 1-12.
- Shiran E., M. S. Mossadegh, and M. Esfandiari. 2012. Symbiotic ants with aphids in Khuzestan province, Isfahan city and its vicinity, Iran. 20th Iranian Plant Protection Congress, 25-28 August 2012.
- Shuang Zhao. 2006. Ant of Guangdong Province (Hymenoptera: Formicidae): A taxonomic study of the ants of Guangdong (Hymenoptera: Formicidae). Master's thesis in Agriculture Insects and Pest Control. Supervisor Fenglong Jia. 115 pages.
- Slipinski P., M. Zmihorski, and W. Czechowski. 2012. Species diversity and nestedness of ant assemblages in an urban environment. Eur. J. Entomol. 109: 197206.
- So, Ha, Seong, Jin, Park, Joon and Byung, and Kim, Jin. 2002. Comparitive Ant Faunas between Seonyudo and Seven other Islands of West Sea in Korea. Korean Journal of Entomology. 32:75-79.
- Sommer F. 1984. Etude de la myrmécofaune de la réserve de la Massane. Laboratoire Arago, Banyuls sur Mer (66), 29 pages.
- Sommer F., and H. Cagniant. 1988. Peuplements de fourmis des Albères Orientales (Pyrénées-Orientales, France) (Première partie). Vie Milieu 38: 189-200.
- Sonnenburg H. 2005. Die Ameisenfauna (Hymenoptera: Formicidae) Niedersachsens und Bremens. Braunschweiger Naturkundliche Schriften 7: 377-441.
- Sonobe R. 1977. Ant fauna of Miyagi prefecture , Japan. Japanese Journal of Ecology 27: 111-116.
- Soulie J. 1962. Fourmis des Hautes-Pyrenees. Bull. Soc. Hist. Nat. Toulouse 97: 35-37.
- Soyer B. 1951. Les fourmis des collines des environs de Marseille. Bull Soc Linn Provence 18: 3-6.
- Steiner F. M., B. Seifert, K. Moder, and B. C. Schlick-Steiner. 2010. A multisource solution for a complex problem in biodiversity research: description of the cryptic ant species Tetramorium alpestre sp. n. (Hymenoptera: Formicidae). Zoologischer Anzeiger 249: 223-254.
- Steiner F. M., S. Schödl, and B. C. Schlick-Steiner. 2002. Liste der Ameisen Österreichs (Hymenoptera: Formicidae), Stand Oktober 2002. Beiträge zur Entomofaunistik 3: 17-25.
- Stukalyuk S. V. 2015. Structure of the ant assemblages (Hymenoptera, Formicidae) in the broad-leaved forests of Kiev. Entomological Review 95(3): 370–387.
- Stukalyuk S. V. 2016. Crematogaster subdentata Mayr, 1877, a potentially invasive species of ant (Hymenoptera, Formicidae) new to the fauna of Crimea. Entomological Review 95(8)
- Stumper R. 1953. Etudes myrmecologiques. XI. Fourmis luxembourgeoises. Bulletin Soc. Nat. luxemb. 57: 122-135.
- Suay-Cano, V. A., A. Tinaut, and J. Selfa. "Las hormigas (Hymenoptera, Formicidae) asociadas a pulgones (Hemiptera, Aphididae) en la provincia de Valencia." Graellsia 58(1) (2002): 21-37.
- Suvak M. 2007. The ants (Hymenoptera: Formicidae) in the stubble-fields of east Slovakia. Acta Zoologica Universitatis Comenianae. 47: 21-33
- Suñer i Escriche, David. "Contribució al coneixement mirmecologic de Gavarres, Montgrí, Guilleríes i la Serralada Transversal." Tesis Doctoral Universida (1991): 577 pp.
- Szujecki A., J. Szyszko, S. Mazur, and S. Perlinski. 1978. A succession of the ants (Formicidae) on afforested arable land and forest soils. Memorabilia Zoologica 29: 183-189.
- Taheri A., J. Reyes-Lopez, and N. Bennas. 2014. Contribution a l'etude de la fauna myrmecologique du parc national de Talassemtane (nord du Maroc): biodiversite, biogeographie et especes indicatrices. Boletín de la Sociedad Entomológica Aragonesa (S.E.A.), 54: 225236.
- Tanaka B., N. Kanie, T. Mano, R. Arita, and A. Shiragane. 1999. The insect fauna between Takabashi bridge and Nomi-kouen Park, margins of the Yahagigawa river. ????? 3: 35-79.
- Tang J., Li S., Huang E., Zhang B. and Chen Y. 1995. Hymenoptera: Formicidae (1). Economic Insect Fauna of China 47: 1-133.
- Terayama M. 1977. Checklist of the known ants of Saitama Prefecture. Insects and nature 12(4): 26-27
- Terayama M. 1978. Ants of Bukozan. Nature and insects 13(4): 32-34.
- Terayama M. 1983. Kagoshima-ken-hondo no ari. Kanagawa-chucho (Journal of the Kanagawa Entomologists Association): 13-24.
- Terayama M. 1992. Structure of ant communities in East Asia. A. Regional differences and species richness. Bulletin of the Bio-geographical Society of Japan 47: 1-31.
- Terayama M., Choi, B.M., Kim, C.H. 1992. A check list of ants from Korea, with taxonomic notes. Bulletin of the Toho Gakuen 7:19-54.
- Terayama M., K. Ogata, and B.M. Choi. 1994. Distribution records of ants in 47 prefectures of Japan. Ari (report of the Myrmecologists Society of Japan) 18: 5-17.
- Terayama M., and K. Murata. 1987. Relation between ant communities and vegetations of Toshima island, the Izu Islands. Bull. Biogeogr. Soc. Japan 42(9): 57-63.
- Terayama M., and K. Murata. 1990. Effects of area and fragmentation of forests for nature conservation: Analysis by ant communities. Bull. Biogeogr. Soc. Japan 45(2): 11-17.
- Teruyama. M. 1988. Ant fauna of Saitama Prefecture, Japan. ARI Reports of the Myrmecologists Society (Japan) 16: 4-13
- Tie Ru, and Xu Shengquan. 2004. Variety and distribution of ants in Northwest China. Journal of Ningxia Agricultural College 25(3): 4-9.
- Tinaut A. 2016. Ants of the Tejeda, Almijara and Alhama Mountains Natural Park (Andalusia, Spain) (Hymenoptera, Formicidae). Boln. Asoc. esp. Ent., 40 (1-2): 125-159.
- Tinaut A., O. Mira, J. M. Vidal, and A. Aguirre-Segura. 2009. The ants of Cabo de Gata (Almería). Faunistics aspects (Hymenoptera, Formicidae). Boln. Asoc. esp. Ent., 33 (1-2): 227-251.
- Tinaut A., and F. J. Ortiz. 1988. Introduccion al conocimiento de las hormigas de la provincia de Almeria (Hymenoptera: Formicidae). Publicaciones del Instituto de Estudios Almerienses. Boletín (Ciencias) 8: 223-231.
- Tinaut, A., J. Jiménez Rojas, and R. Pascual. "Estudio de la mirmecofauna de los bosques de Quercus Linneo 1753 de la provincia de Granada." Ecología 8 (1995): 429-438.
- Tinaut, A. "Contribución al estudio de los formícidos de la región del estrecho de Gibraltar y su interés biogeográfico." Graellsia 45 (1989): 19-29.
- Tohme G. 1996. Formicidae. Etude de la diversité biologique n° 4 . Ministère de lAgriculture à Beyrouth (Eds.). P85-87.
- Tohme G., and H. Tohme. 2014. Nouvelles liste des especes de fourmis du Liban (Hymenoptera, Formicoidea). Lebanese Science Journal 15(1): 133-141.
- Urbieta Balado A., A. Palanca Soler, C. Castan Lanaspa, and X. Espadaler. 1984. Ants in the supraforestal pastures of the Tendenera massif. Documents dEcologie Pyrénéenne III-IV: 223-224.
- Vesni? A. 2011: Revidirani sistematski prijegled mrava Bosne i Hercegovine. Unutar : S. Lelo (urednik), Fauna Bosne i Hercegovine Biosistematski pregledi. 7. izmijenjeno i popravljeno interno izdanje Udruenja za inventarizaciju i zatitu ivotinja, Ilija, Kanton Sarajevo, pp: 205-207.
- Vogrin, V. "Prilog fauni Hymenoptera - Aculeata Jugoslavije." Zast. Bilja 31(suppl.) (1955): 1-74.
- Wagner H. C., W. Arthofer, B. Seifert, C. Muster, F. M. Steiner, and B. C. Schlick-Steiner. 2017. Light at the end of the tunnel: Integrative taxonomy delimits cryptic species in the Tetramorium caespitum complex (Hymenoptera: Formicidae). Myrmecological News 25: 95-129.
- Wang 2008. Ant species diversity on Shangqiu forest park, Henan province. Sichuan Journal of Zoology 27(6): 1041-1044.
- Wang Lu-ling. 2008. Ant species diversity on Shangqiu forest Park, Henan province. Sichuan Journal of Zoology 27(6): 1041-1044.
- Wang Minsheng, Xiao Gangrou, and Wu Jian. 1988. Taxonomic studies on the ant genus Tetramorium Mayr in China (Hymenoptera: Formicidae). Forest Research 1(3): 264-274.
- Wang S. Z. et al. 2011. Investigation on ant species for garden tree in Chengdu. Journal of Anhui Agri. Sci. 39(9): 5045-5046.
- Wang Wei et al. 2006. Survey of ant species in three nature reserves in Eastern Hubei province. Journal of Anhui Agri. Sci. 34(13): 3131-3132.
- Wegnez P. 2014. Premières captures de Lasius distinguendus Emery, 1916 et de Temnothorax albipennis (Curtis, 1854) au Grand-Duché de Luxembourg (Hymenoptera : Formicidae). Bulletin de la Société royale belge d’Entomologie 150 (2014) : 168-171.
- Wegnez P. 2017. Découverte de Myrmica lobicornis Nylander, 1846 et Lasius jensi Seifert, 1982, deux nouvelles espèces pour le Grand-Duché de Luxembourg (Hymenoptera: Formicidae). Bulletin de la Société royale belge d’Entomologie 153: 46-49.
- Wegnez P. 2018. Premières decouvertes de Myrmica bibikoffi Kutter, 1963 et de Ponera testacea Emery, 1895, au Luxembourg (Hymenoptera: Formicidae)Bulletin de la Société royale belge d’Entomologie 154: 263–272.
- Wegnez P., D. Ignace, E. Lommelen, M. Hardy, J. Bogaert, and C. Nilsson. 2015. Redécouverte de Teleutomyrmex schneideriKutter, 1950 dans les Alpes françaises (Hymenoptera: Formicidae). Bulletin de la Société royale belge d’Entomologie 151: 52-57.
- Wegnez P., and A. Ronk. 2017. Découverte de Camponotus herculeanus (Linnaeus, 1758) et signalement de quelques autres espèces rares de fourmis au Luxembourg (Hymenoptera : Formicidae). Bulletin de la Société des naturalistes luxembourgeois 119 : 153–159.
- Wegnez P., and M. Fichaux. 2015. Liste actualisee des especes de fourmis repertoriees au Grand-Duche de Luxembourg (Hymenoptera: Formicidae). Bulletin de la Société royale belge d’Entomologie 151: 150-165
- Wetterer J. K., X. Espadaler, A. L. Wetterer, and S. G. M. Cabral. 2004. Native and exotic ants of the Azores (Hymenoptera: Formicidae). Sociobiology 44: 1-20.
- Wetterer J. K., X. Espadaler, A.L. Wetterer, and S.G.M. Cabral. 2004. Native and exotic ants of the Azores (Hymenoptera: Formicidae). Sociobiology 44: 1-20.
- Wetterer, James K. and Wetterer, Andrea L. 2004. Ants (Hymenoptera: Formicidae) of Bermuda. Florida Entomologist 87(2): 212-221.
- Wetterer, James K., Wetterer, Andrea L., and Cabral, Susana G. M. 2004. Native and Exotic Ants of the Azores (Hymenoptera: Formicidae). Sociobiology. 44(1):1-20.
- Wheeler W. M. 1906. The ants of Japan. Bulletin of the American Museum of Natural History 22: 301-328.
- Wheeler W. M. 1921. Chinese ants. Bulletin of the Museum of Comparative Zoology 64: 529-547.
- Wheeler, W. M. 1936. Ants from the Society, Austral, Tuamotu and Mangareva Islands. Occasional Papers of the Bernice Pauahi Bishop Museum 12(18):3-17.
- Wheeler, William Morton. 1906. The Ants of the Bermudas. Bulletin American Museum of Natural History. 22:347-352.
- Wheeler, William Morton. 1909. Ants from the Azores. Bulletin American Museum of Natural History. 24:169-170.
- Wheeler, William Morton. 1936. Ants from the Society, Austral, Tuamotu, and Mangareva Islands. Bernice P. Bishop Museum Occasional Papers 7.18.
- Wiezick M. 2008. Ant communities (Hymenoptera: Formicidae) of forest-steppe habitats at southern and South-Western slopes of Plesivecka a Planina plateau at Slovensky Kras Karst. Natura Carpatica 49: 85-94.
- Wiezik M. 2006. Ants (Hymenoptera: Formiciadae) of the Sitno National Nature Reserve (tiavnické vrchy Protected Landscape Area). Naturae Tutela 10: 73-77.
- Wiezik M., A. Wiezikova, and M. Svitok. 2010. Effects of secondary succession in abandoned grassland on the activity of ground-foraging ant assemblages (Hymenoptera: Formicidae). Acta Soc. Zool. Bohem. 74: 153-160
- Wiezik M., A. Wiezikova, and M. Svitok. 2011. Vegetation structure, ecological stability, and low-disturbance regime of abandoned dry grasslands support specific ant assemblages in Central Slovakia. Tuexenia 31: 301315.
- Wieziková A. 2008. Epigaeic activity of ants (Hymenoptera: Formicidae) at hemerobic gradient: arable land green forage permanent grassland. Acta Facultatis Ecologiae 18: 69-76.
- Wilson EO, Taylor RW. 1967. The ants of Polynesia. Pacific Insects Monograph 14:1-109.
- Wu J. and Wang C. 1995. The ants of China. China Forestry Publishing House, Beijing. 214 pp.
- Wu W. 2010. The taxonomic and fauna study on the Formicidae of Liaoning Province (Insecta: Hymenoptera). Master's Thesis Northeast Normal University, 75 pages.
- Xin M. et al. 2011. Community structure of Hymenoptera fauna in Helan Mountain of Ningxia. Journal of Anhui Agri. Sci. 39(25): 15339-15341.
- Xin M., Y. Ma, and D. He. 2011. Fauna composition of Formicidae in Ningxia. Journal of Ningxia University (Natural Science Edition) 32(4): 403-412.
- Yap H. H., and C. Y. Lee. 1994. A preliminary study on the species composition of household ants on Penang island, Malaysia. Journal of Bioscience 5(1-2): 64-66.
- Yarrow, I.H.H. 1967. On the Formicidae of the Azores. Blotim Do Museu Municipal Do Funchal. XXI(97):24-32.
- Yoshimoto T., and S. Yamane. 1990. Food habits of some ant species inhabiting the Taisho lava of the Sakura-jima volcano, Japan (Hymenoptera, Formicidae). Reports of the Faculty of Science Kagoshima University 23: 9-22.
- Zhang W. and Zheng Z. 2002. Studies of ant (Hymenoptera: Formicidae) fauna in Sichuan Province. Entomotaxonomia 24(3): 216-222.
- Zhou S.-Y. 2001. Ants of Guangxi. Guangxi Normal University Press, Guilin, China, Guilin, China. 255 pp.
- Zhu C., K. Chi, X. Ye, H. Zhou, and C. Lio. 2010. Community structure and species diversity of ants in Xuzhou, Jiansu. Journal of Forestry Science and Technology 37(2): 7-10.
- Zhu Xun. 2002. Study on flora of ants in Hunan province. Journal of Hunan Environment-Biological Polytechnic 8(3): 172-174.
- Zhuytszyuan D. 2016. The ants (Hymenoptera, Formicidae) Nizhne-Svirsky reserve and their environmental features. Master's thesis Saint Petersburg State University.
- Zimmermann, S. "Beitrag zur Kenntnis der Ameisenfaunader Quarnerischen Inseln." Verhandlungen der Zoologisch-Botanisch Gesellschaft in Wien 80 (1930): 45-66.
- Zryanin V. A., and T. A. Zryanina. 2007. New data on the ant fauna Hymenoptera, Formicidae in the middle Volga River Basin. Uspekhi Sovremennoi Biologii 127(2): 226-240.
- de Haro, Andrés, C. A. Collingwood, and P. Comín del Río. "Prospección mirmecológica por Ibiza y Formentera (Baleares)." Orsis 2 (1986): 115-120.
- de Haro, Andrés, and C. A. Collingwood. "Prospección mirmecológica en la Cordillera Ibérica." Orsis 6 (1991): 129-126.
- de Haro, Andrés. "Contribución al conocimiento de los formícidos del cabo de Creus (Gerona)." Pirineos 117 (1982): 49-57.
- Pages using DynamicPageList3 parser function
- Photo Gallery
- North temperate
- North subtropical
- FlightMonth
- Ant Associate
- Host of Strongylognathus karawajewi
- Host of Tetramorium atratulum
- Host of Tetramorium inquilinum
- Host of Strongylognathus testaceus
- Host of Strongylognathus christophi
- Host of Strongylognathus huberi
- Host of Strongylognathus italicus
- Host of Strongylognathus kratochvili
- Host of Formica fusca
- Host of Formicoxenus nitidulus
- Host of Lasius flavus
- Aphid Associate
- Host of Aphis fabae
- Host of Aphis farinosa
- Host of Aphis gossypii
- Host of Aphis salviae
- Host of Aphis spiraecola
- Host of Brachycaudus cardui
- Host of Macrosiphoniella sanborni
- Host of Myzus lythri
- Nematode Associate
- Host of Mermithidae (unspecified "Mermix")
- Host of Oscheius dolichura
- Mite Associate
- Host of Petalomium nataliae
- Host of Scutacarus longisetus
- Cestode Associate
- Host of Raillietina caucasica
- Host of Raillietina circumvalata
- Host of Raillietina echinobothrida
- Host of Raillietina fedjuschini
- Host of Raillietina tetragona
- Fungus Associate
- Host of Myrmicinosporidium durum
- Host of Tarichium sp.
- Karyotype
- Species
- Extant species
- Formicidae
- Myrmicinae
- Crematogastrini
- Tetramorium
- Tetramorium caespitum
- Myrmicinae species
- Crematogastrini species
- Tetramorium species
- Ssr