Tapinoma melanocephalum
| Tapinoma melanocephalum | |
|---|---|

| |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Arthropoda |
| Class: | Insecta |
| Order: | Hymenoptera |
| Family: | Formicidae |
| Subfamily: | Dolichoderinae |
| Genus: | Tapinoma |
| Species: | T. melanocephalum |
| Binomial name | |
| Tapinoma melanocephalum (Fabricius, 1793)
| |
| Synonyms | |
| |


| Common Name | |
|---|---|
| Ghost Ant | |
| Language: | English |
| Awate-konuka-ari | |
| Language: | Japanese |
This is a widespread tropical tramp species, introduced throughout the world. Nests are found in the soil, under stones, fallen trees, tree bark and in other crevices. Its native range is unknown. It is ubiquitous in non-air conditioned dwellings anywhere in the lowland tropics. Regardless where you are, if you are sitting at a table with a sugar dispenser you are likely to see workers of T. melanocephalum running about on the surface. They always seem to be able to find their way into the sugar container, and sugar on tropical tables always contains some non-negligible fraction of T. melanocephalum workers. When you put a spoonful of sugar in your drink, you can judge the level of contamination by how many workers are left floating on the surface.
In quantitative biodiversity surveys, this species often has to be excluded from data analysis because the laboratory where samples are processed contains T. melanocephalum as a pest, and contamination of samples occurs.
Although most often found in houses, they can also move out into surrounding vegetation in highly disturbed and highly insolated habitats, opportunistically nesting in small plant cavities. Nests readily relocate, and overnight they can move into a shoe or an umbrella left on a porch. (Jack Longino)
| At a Glance | • Highly invasive • Polygynous • Supercolonies |
Photo Gallery

{kind=link}
Identification
Distinctively bicoloured, head brown, alitrunk and gaster pale yellowish with variable brown patches; legs and antennae very pale. Prothorax laterally compressed, broadening anteriorly. Length: 1.5-2 mm. (Collingwood 1979)
Keys including this Species
- Key to US Tapinoma species
- Dolichoderinae of the southwestern Australian Botanical Province
- Key to workers of the Socotra Archipelago, Yemen
- Key to Tapinoma of Hispaniola
- Key to Palaearctic Tapinoma
- Key to queens of Nearctic Tapinoma
- Key to males of Nearctic Tapinoma
Distribution
Tapinoma melanocephalum has one of the widest distributions known for any ant species. It has spread across the Old World and New World in both the northern and southern hemispheres. It is tropical and subtropical; at latitudes greater than 30°, it is largely restricted to living inside buildings. The only ants with broader known distributions than T. melanocephalum are the longhorn crazy ant, Paratrechina longicornis, and the pharaoh ant, Monomorium pharaonis. I expect that T. melanocephalum will eventually spread to the disturbed lowlands of virtually every tropical country. This species also appears to be an increasingly common indoor pest in temperate areas, and some-day may be found in every major city of the world. Because T. melanocephalum can live anywhere that humans live, there is no limit to the latitude where indoor populations of this species may be found. (Wetterer 2009)
Latitudinal Distribution Pattern
Latitudinal Range: 32.812778° to -9.233°.
| North Temperate |
North Subtropical |
Tropical | South Subtropical |
South Temperate |
- Source: AntMaps
Distribution based on Regional Taxon Lists
Afrotropical Region: Benin, Cape Verde, Comoros, Gambia, Guinea, Nigeria, Saint Helena, Senegal, Socotra Archipelago, United Arab Emirates, Yemen.
Australasian Region: Australia, New Caledonia, Norfolk Island.
Indo-Australian Region: Borneo, Cook Islands, Fiji, French Polynesia, Guam, Hawaii, Indonesia (type locality), Krakatau Islands, Malaysia, New Guinea, Niue, Philippines, Samoa, Singapore, Solomon Islands, Tokelau, Tonga, Vanuatu (type locality), Wallis and Futuna Islands.
Malagasy Region: Madagascar, Mauritius, Mayotte, Réunion, Seychelles.
Nearctic Region: Canada, United States.
Neotropical Region: Anguilla, Bahamas, Barbados, Belize, Brazil, Cayman Islands, Colombia (type locality), Costa Rica (type locality), Cuba, Dominican Republic, Ecuador, French Guiana, Galapagos Islands, Greater Antilles, Grenada, Guadeloupe, Guatemala, Guyana, Haiti, Honduras, Lesser Antilles, Mexico, Netherlands Antilles, Paraguay, Puerto Rico, Suriname, Trinidad and Tobago.
Oriental Region: Bangladesh, Cambodia, India (type locality), Laos, Maldives, Nicobar Island, Pakistan, Sri Lanka, Thailand.
Palaearctic Region: Afghanistan, Belgium, Canary Islands, China, Czechia, Finland, Hungary, Iberian Peninsula, Iceland, Italy, Japan, Oman, Republic of Korea, Romania, Russian Federation, Spain, Ukraine, United Kingdom of Great Britain and Northern Ireland.
Distribution based on AntMaps
Distribution based on AntWeb specimens
Check data from AntWeb
Countries Occupied
| Number of countries occupied by this species based on AntWiki Regional Taxon Lists. In general, fewer countries occupied indicates a narrower range, while more countries indicates a more widespread species. |

|
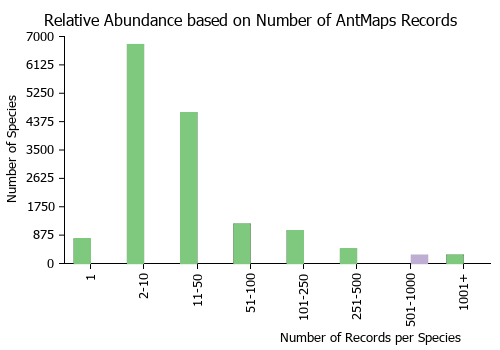
Estimated Abundance
| Relative abundance based on number of AntMaps records per species (this species within the purple bar). Fewer records (to the left) indicates a less abundant/encountered species while more records (to the right) indicates more abundant/encountered species. |
|
Biology
San Cristóbal, República Dominicana. Video by Judá Isaí Martínez Uribe.
Wetterer (2009) - "Tapinoma melanocephalum is a ubiquitous pest through much of the tropics and subtropics. As Longino (2006) aptly wrote: "regardless of whether you are in Guinea, New Guinea, or Guyana, if you are sitting at a table with a sugar dispenser you are likely to see workers of T. melanocephalum running about on the surface." Workers are small (~ 1.5 mm) and their pale legs and abdomens often blend into the background, making them difficult to see. Their dark brown heads and thoraces often look like hovering specks, unrecognizable as ants. The barely visible "ghostly" appearance of T. melanocephalum no doubt explains its common name.
Tapinoma melanocephalum is particularly noted as a pest in homes, restaurants, hospitals, and greenhouses. For example, T. melanocephalum was the most common house ant in Brazil (e.g., Kamura et al. 2007) and on Panang Island, Malaysia (Lee et al. 2002), and it was the most fre-quently reported pest ant in greater Miami (Klotz et al. 1995). Wheeler (1907) noted that T. melanocephalum had been "introduced into well-heated buildings in New York," and observed workers in the winter "visiting the sugar bowls on the tables of the Columbia University commons." Karawajew (1926) reported T. melanocephalum from hotel tables both on Ambon Island, Indonesia and in Vladivostok, Russia. James Trager (pers. comm.) observed T. melanocephalum inside the Sachs Butterfly House in Chesterfield, Missouri, beginning shortly after it opened in 1995. Tapinoma melanocephalum also has been a long term problem in the greenhouses of the Dow Estate in Midland, Michigan, where fumigation is incompatible with a butterfly exhibit (J. LaForest, pers. comm.).
Although T. melanocephalum does not sting and its bite is not very painful (pers. obs.), it can be a serious pest. For example, Fowler et al. (1993) and Moreira et al. (2005) found that T. melanocephalum was the most prevalent ant in hospitals in Brazil, and had the highest potential as a vector for pathogens. In fact, Moreira et al. (2005) found at least 14 different types of bacteria on T. melanocephalum workers collected in the hospitals, including antibiotic resistant strains. Tapinoma melanocephalum also can be an agricultural pest because it tends populations of phloem feeding Hemiptera, such as mealybugs, scale insects, and aphids, for their honeydew (Venkataramaiah and Rehman 1989). Hemiptera cause damage by sapping plants of nutrients and increasing the occurrence of diseases, including viral and fungal infections. Fowler et al. (1994) concluded that T. melanocephalum had a great impact on other ant species on banana plantations in Brazil. Under some conditions, T. melanocephalum may be beneficial as a biocontrol agent. For example, Osborne et al. (1995) found that in greenhouses of central Florida, naturally occurring populations of T. melanocephalum were important in controlling a serious plant pest, the two-spotted spider mite, Tetranychus urticae Koch, 1836.
Whereas T. melanocephalum is most prevalent in disturbed habitats, it has also been reported from natural reserves around the world, including the Baiyer River Wildlife Sanctuary in Papua-New Guinea (Whalen and Mackay 1988), Reserva Biológica de Una in Brazil (Delabie et al. 1997), primary rainforest in Chocó, Colombia (Armbrecht et al. 2001), and on many Galapagos islands (Causten et al. 2006). In the Wapoga River Area of Irian Jaya, Indonesia, Snelling (2000) found that T. melanocephalum "had successfully invaded forest areas where they apparently became quickly assimilated into the native mix; their presence did not seem to adversely affect other ant species there. This apparently benign situation probably would be reversed in the presence of continued disturbance of the habitat."
Tapinoma melanocephalum colonies are polygynous and frequently relocate their nests. In a matter of hours, colonies can move into cargo, which is then shipped worldwide. In fact, Lester (2005) reported that T. melanocephalum was the second most common ant species (after Pheidole megacephala) intercepted in freight entering New Zealand between 1997 and 2002. Weber (1939) reported collecting T. melanocephalum from riverboats in Venezuela and Guyana and on a ship between Haiti and New York. Tapinoma melanocephalum was found on a train in France, traveling from Paris to Rennes (X. Espadaler, pers. comm.). In Le Carbet, Martinique, a T. melanocephalum colony moved overnight into luggage packed for departure, requiring me to shake out everything before repacking."
Klimes and Okrouhlik (2015) - One of the most frequently recorded pests in hospitals in the tropics . . . In buildings, it is usually seen in kitchen corridors and bathrooms . . . Despite their diminutive size, workers are able to use their mandibles to penetrate even the thick plastic of sugar packages.
Although this species has been listed as supercolonial (Debout et al., 2007; Eyer & Vargo, 2021), genetic evidence of this is lacking.
Regional Notes
Espadaler (2007) - Canary Islands: Running workers were detected in irrigated gardens next to recently built bungalows. This is the first record for El Hierro of this well known tramp species.
Klimes and Okrouhlik (2015) report on a indoor infestation in an apartment complex in South Bohemia, Czech Republic. This is the first time Tapinoma melanocephalum has been recorded from the country.
This species is common in south Florida, to Volusia county. Almost always in disturbed areas, in many sites seems confined to areas around buildings. Nests are in bark mulch, under objects on the ground, and under loose bark and the bases of palm fronds. Pest status: can enter buildings through screens and small cracks, becoming a general annoyance. First published Florida record: Smith 1930. (Deyrup, Davis & Cover, 2000.)
Sharaf et al. (2017) - Yemen: The nesting and foraging habitats of this species are diverse. Several specimens were found foraging on a tree located on a mountainside with nearby stream drainage. The area had moist soils with high plant diversity, especially Adiantum capillus-veneris L. (Pteridaceae), which was abundant. The area was dominated by the ponerine ant, Brachyponera sennaarensis (Mayr, 1862). A nest series was collected from an area with moist soil and dense grasses. The area was rich in decayed animal faeces. Another nest was collected by sifting the leaf litter next to a stream and small pool with many scattered date palm (Phoenix dactylifera L.) trees. Another nest series was found under the bark of a recently cut dragon blood tree, Dracaena cinnabari Balf.f. (Asparagaceae) and associated with curculionid beetles. This invasive species has a broad range of habitat preference worldwide (Wetterer 2009a), including pre-existing cavities made of plant materials, and in larders and pantries, and nesting in walls and potted plants indoors (Ellison et al. 2012).
Foraging/Diet
An experimental study by Zhou et al. (2015) tested effects of ant tending, host plants, parasitism and their interactions on mealybugs on two host plant species. The plants, tomato (Solanum lycopersicun) and cotton (Gossypium hirsutum), were grown in a greenhouse at an agricultural field station near Guangzhou, China. The ants (Tapinoma melanocephalum), hemipterans (Phenacoccus solenopsis Tinsley) and parasitoid (Aenasius bambawalei Hayat) were collected from local habitats, i.e., are naturally sourced and locally occurring species that are known to interact with one another. They found that mealybug benefits from ant attendance by T. melanocephalum did differ by host plant. The ant tending was more intense on tomato versus cotton plants. The mealybugs showed both a higher growth rate with higher ant attendance and a decrease in parasitism rate. In sum, they found the tomato plants likely produced higher quality honeydew, stimulating ant attendance, and this in turn provided positive benefits to the hemiptera.
Cheng et al. (2019), studying this species in Guangzhou China, found a number of differences in workers sampled from populations of T. melanocephalum from areas with Solenopsis invicta versus areas where this other ant is not present. For many analyses and experiments that were part of this study, Tapinoma melanocephalum workers were sampled from numerous colonies within each of three sites with S. invicta and three sites without. It appears competition between these two invasive species alters T. melancephalum aggressiveness (less aggressive when S. invicta is present), diet (stable isotype analysis suggests a switch from a carbohydrate-rich to a protein-rich diet when S. invicta is present), microbial bacteria symbionts, and other bacteria (the relative abundance of Wolbachia, Lactobacillales and Enterobacteriaceae). Being less aggressive potentially reduces harmful interactions between T melanocephalum and S. invicta. The change in diet, and a shift away from nectar and honeydew sources that are likely exploited by S. invicta, is an apparent adjustment to avoid competition (or is simply a result of competing poorly). This dietary change in turn putatively alters the workers microbial symbionts and bacteria.
Association with Other Organisms
 Explore: Show all Associate data or Search these data. See also a list of all data tables or learn how data is managed.
Explore: Show all Associate data or Search these data. See also a list of all data tables or learn how data is managed.
Other Insects
This ant has been observed tending larvae of Lampides boeticus (Obregon et al. 2015).
This species is known to tend the following aphids in Pakistan (Siddiqui et al., 2019; Gull-E-Fareen et al., 2020): Aphis eugeniae, Aphis fabae, Aphis fabae solanella, Aphis gossypii, Aphis pomi, Greenidea (Greenidea) decaspermi, Greenidea (Greenidea) ficicola, Greenidea (Trichosiphum) formosana, Hysteroneura setariae, Macrosiphoniella sanborni, Myzus persicae, Ovatus crataegarius, Pentalonia nigronervosa, Tinocallis kahawaluokalani and Tuberolachnus salignus.
Chemistry
Shi et al. (2017) - Abstract Volatile compounds in Tapinoma melanocephalum (Fabricius) (Hymenoptera: Formicidae, Dolichoderinae) workers were analyzed using gas chromatography–mass spectrometry and electroantennogram responses of workers to these compounds were investigated using coupled gas chromatography–electroantennographic detection. Among 18 detected compounds, six elicited electroantennogram response, including 6-methyl-5-hepten-2-one, 4 iridodials, and (Z)-9-nonadecene. (Z)-9-Nonadecene and iridodials were identified the first time in T. melanocephalum. (Z)-9-Nonadecene is a major component in mandibular glands and iridodials are produced in pygidial glands. In contrast to previous report, actinidine was not found in pygidial glands. Behavioral response of workers to the synthetic (Z)-9-nonadecene was investigated. (Z)-9-nonadecene is an attractant to T. melanocephalum. It also affected their locomotion patterns.
Life History Traits
- Queen number: polygynous (Holldobler & Wilson, 1977)
Castes
Worker
                   
| |
| . | |
Additional worker images can be found here
Queen
Images from AntWeb
   
| |
| Queen (alate/dealate). Specimen code casent0444732. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by CAS, San Francisco, CA, USA. |
   
| |
| Queen (alate/dealate). Specimen code casent0173214. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by CDRS, Galapagos, Ecuador. |
   
| |
| Queen (alate/dealate). Specimen code casent0125327. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by CAS, San Francisco, CA, USA. |
Male
    
| |
| . | |
Nomenclature
The following information is derived from Barry Bolton's Online Catalogue of the Ants of the World.
- melanocephalum. Formica melanocephala Fabricius, 1793: 353 (w.) FRENCH GUIANA.
- [Misspelled as melanorephalum by Santschi, 1924c: 112.]
- Emery, 1887a: 249 (m.); Forel, 1891b: 102 (q.); Wheeler, G.C. & Wheeler, J. 1951: 197 (l.); Crozier, 1970: 119 (k.).
- [Note: neotype w. designated by Guerrero, 2018: 499. Colombia: Magdalena. Guerrero, 2018: 499, also nominates “paraneotypes”: under ICZN rules no such category exists.]
- Combination in Lasius: Fabricius, 1804: 417.
- Combination in Myrmica: Lepeletier de Saint-Fargeau, 1835: 185.
- Combination in Micromyrma: Roger, 1862b: 258.
- Combination in Tapinoma: Mayr, 1862: 651; Dalla Torre, 1893: 165.
- Combination in Tapinoma (Micromyrma): Santschi, 1928e: 475.
- Status as species: Latreille, 1802c: 269; Fabricius, 1804: 417; Lepeletier de Saint-Fargeau, 1835: 185; Smith, F. 1858b: 46; Roger, 1862b: 258; Roger, 1863b: 14; Mayr, 1863: 455; Mayr, 1876: 83; Forel, 1881: 3; Mayr, 1884: 31; Mayr, 1886c: 359; Emery, 1887a: 249; Forel, 1891b: 101 (redescription); André, 1892b: 51; Forel, 1893g: 352; André, 1893b: 152; Dalla Torre, 1893: 164; Emery, 1893d: 89; Emery, 1893e: 194; Emery, 1893f: 249; Emery, 1893g: 267; Forel, 1895a: 49; Forel, 1895b: 109; Forel, 1895e: 472; Emery, 1895f: 337; Pergande, 1896: 870; Mayr, 1897: 432; Emery, 1897d: 573; Forel, 1899a: 120; Forel, 1899c: 101; Emery, 1900d: 696; Emery, 1901f: 121; Forel, 1902h: 476; Rothney, 1903: 98; Bingham, 1903: 304; Wheeler, W.M. 1905b: 131; Emery, 1906c: 177; Wheeler, W.M. 1907a: 275; Forel, 1907e: 9; Wheeler, W.M. 1908a: 150; Forel, 1908e: 64; Wheeler, W.M. 1909c: 272; Wheeler, W.M. 1909d: 336, 342; Yano, 1910: 421; Wheeler, W.M. 1911a: 29; Wheeler, W.M. 1912a: 46; Forel, 1912k: 164; Emery, 1913a: 41; Forel, 1913f: 197; Forel, 1913k: 93; Wheeler, W.M. 1913b: 498; Wheeler, W.M. 1913d: 242; Wheeler, W.M. 1913e: 237; Santschi, 1914d: 377; Emery, 1914f: 420; Donisthorpe, 1915d: 342; Viehmeyer, 1916a: 142; Mann, 1916: 473; Wheeler, W.M. 1916d: 330; Crawley, 1916b: 375; Donisthorpe, 1918b: 167; Wheeler, W.M. 1919c: 275; Wheeler, W.M. 1919e: 100; Santschi, 1920h: 172; Mann, 1921: 473; Wheeler, W.M. 1922a: 924, 1034; Wheeler, W.M. 1922c: 15; Borgmeier, 1923: 101; Wheeler, W.M. 1923b: 4; Wheeler, W.M. 1924a: 110; Crawley, 1924: 403; Santschi, 1924c: 112; Mann, 1925b: 6; Mukerjee & Ribeiro, 1925: 207; Karavaiev, 1926d: 441; Stärcke, 1926: 118 (in key); Donisthorpe, 1927b: 395; Wheeler, W.M. 1927d: 7; Wheeler, W.M. 1927h: 99; Wheeler, W.M. 1928c: 30; Cheesman & Crawley, 1928: 522; Santschi, 1928c: 70; Wheeler, W.M. 1929f: 8; Wheeler, W.M. 1929g: 57; Karavaiev, 1930a: 213; Menozzi, 1930b: 114; Menozzi, 1930d: 328; Menozzi & Russo, 1930: 165; Smith, M.R. 1930a: 5; Wheeler, W.M. 1930h: 73; Donisthorpe, 1932c: 449; Wheeler, W.M. 1932a: 13; Wheeler, W.M. 1932d: 17; Wheeler, W.M. 1934a: 178; Wheeler, W.M. 1934h: 16; Donisthorpe, 1935: 634; Menozzi, 1935b: 200; Wheeler, W.M. 1935g: 36; Wheeler, W.M. 1936b: 204; Wheeler, W.M. 1937a: 23; Smith, M.R. 1937: 861; Teranishi, 1940: 59; Yasumatsu, 1940a: 68; Donisthorpe, 1941h: 60; Santschi, 1941: 277; Wheeler, W.M. 1942: 252; Menozzi, 1942: 178; Eidmann, 1944: 459, 469; Donisthorpe, 1948g: 139; Weber, 1948b: 85; Creighton, 1950a: 352; Smith, M.R. 1951a: 838; Azuma, 1951: 88; Chapman & Capco, 1951: 192; Smith, M.R. 1954c: 10; Kempf, 1961b: 520; Wilson, 1962c: 18; Linsley & Usinger, 1966: 175; Baltazar, 1966: 263; Wilson & Taylor, 1967: 80; Taylor, 1967b: 1094; Kempf, 1972a: 247; Alayo, 1974: 24 (in key); Smith, D.R. 1979: 1421; Collingwood, 1979: 36; Collingwood, 1985: 243; Deyrup, et al. 1989: 99; Wu, J. & Wang, 1992: 1311; Shattuck, 1994: 147; Douwes, 1995: 91; Bolton, 1995b: 400; Wu, J. & Wang, 1995: 116; Dorow, 1996a: 82; Tiwari, 1999: 83; Deyrup, et al. 2000: 300; Mathew & Tiwari, 2000: 342; Zhou, 2001b: 150; Blard, et al. 2003: 133; Deyrup, 2003: 47; Imai, et al. 2003: 84; Collingwood, et al. 2004: 486; Ghosh, et al. 2005: 10; Wild, 2007b: 25; Clouse, 2007b: 206; Don, 2007: 191; Framenau & Thomas, 2008: 59; Terayama, 2009: 199 (in key); Heterick, 2009: 58; Boer, 2010: 16; Collingwood, et al. 2011: 415; Csösz, et al. 2011: 58; Ellison, et al. 2012: 105; Sarnat & Economo, 2012: 37; Sharaf, Aldawood & El Hawagry, 2012a: 40 (in key); Borowiec, L. 2014: 168 (see note in bibliography); Wetterer, et al. 2016: 20; Radchenko, 2016: 92; Sharaf, Fisher, et al. 2017: 5; Deyrup, 2017: 176; Sharaf, Fisher, et al. 2018: 3; Guerrero, 2018: 499 (redescription).
- Senior synonym of australis: Wilson & Taylor, 1967: 80; Shattuck, 1994: 148; Bolton, 1995b: 399; Radchenko, 2016: 92; Guerrero, 2018: 500.
- Senior synonym of coronatum: Guerrero, 2018: 500.
- Senior synonym of familiaris: Dalla Torre, 1893: 165; Forel, 1895b: 109; Forel, 1895e: 472; Pergande, 1896: 870; Forel, 1899c: 101; Wheeler, W.M. 1908a: 150; Emery, 1913a: 41; Wheeler, W.M. 1919c: 276; Wheeler, W.M. 1919e: 100; Wheeler, W.M. 1922a: 925; Borgmeier, 1923: 101; Creighton, 1950a: 352; Smith, M.R. 1954c: 10; Guerrero, 2018: 499.
- Senior synonym of luffae: Guerrero, 2018: 500.
- Senior synonym of malesiana: Guerrero, 2018: 500.
- Senior synonym of nana: Emery, 1892b: 166; Dalla Torre, 1893: 165; Forel, 1895b: 109; Forel, 1895e: 472; Pergande, 1896: 870; Forel, 1899c: 101; Wheeler, W.M. 1908a: 150; Emery, 1913a: 41; Wheeler, W.M. 1919c: 275; Wheeler, W.M. 1919e: 100; Wheeler, W.M. 1922a: 925; Borgmeier, 1923: 101; Creighton, 1950a: 352; Smith, M.R. 1954c: 10; Radchenko, 2016: 92; Guerrero, 2018: 499.
- Senior synonym of pellucida: Mayr, 1886c: 359; Forel, 1891b: 102; Dalla Torre, 1893: 165; Forel, 1895b: 109; Forel, 1895e: 472; Pergande, 1896: 870; Forel, 1899c: 101; Wheeler, W.M. 1908a: 150; Emery, 1913a: 41; Wheeler, W.M. 1919c: 275; Wheeler, W.M. 1919e: 100; Wheeler, W.M. 1922a: 925; Borgmeier, 1923: 101; Creighton, 1950a: 352; Smith, M.R. 1954c: 10; Radchenko, 2016: 92; Guerrero, 2018: 499.
- Senior synonym of wallacei: unavailable name (proposed in synonymy), and also unnecessary (second) replacement name: Guerrero, 2018: 499.
- australis. Tapinoma (Micromyrma) melanocephalum var. australis Santschi, 1928a: 53 (w.) VANUATU (date of publication 25.ii.1928), SAMOA (Upolu I., Savai’i I.), AMERICAN SAMOA (Tutuila I.).
- [Also described as new, Tapinoma (Micromyrma) melanocephalum var. australe Santschi, 1928e: 475 (date of publication 30.viii.1928).]
- Subspecies of melanocephalum: Wheeler, W.M. 1932c: 161; Wheeler, W.M. 1932d: 18; Wheeler, W.M. 1933f: 143; Wheeler, W.M. 1935g: 36; Wheeler, W.M. 1936f: 14.
- Junior synonym of melanocephalum: Wilson & Taylor, 1967: 80; Shattuck, 1994: 148; Bolton, 1995b: 399.
- coronatum. Tapinoma melanocephalum var. coronatum Forel, 1908b: 62 (w.) COSTA RICA.
- Subspecies of melanocephalum: Emery, 1913a: 41; Kempf, 1972a: 247; Shattuck, 1994: 148; Bolton, 1995b: 400.
- Junior synonym of melanocephalum: Guerrero, 2018: 500.
- familiaris. Formica familiaris Smith, F. 1860b: 96 (w.) INDONESIA (Bacan I.).
- [Junior primary homonym of Formica familiaris Smith, F. 1860a: 68.]
- Status as species: Mayr, 1863: 414.
- Replacement name: Formica domestica Smith, F. 1871a: 308.
- Senior synonym of Formica wallacei Guerrero, 2018: 499. [Unavailable name (proposed in synonymy).] Unnecessary (second) replacement name for Formica familiaris Smith, F. 1860b: 96; junior synonym of Formica melanocephala Fabricius, 1793: 353: Guerrero, 2018: 499.
- Junior synonym of melanocephalum: Forel, 1895e: 472; Pergande, 1896: 870; Forel, 1899c: 101; Guerrero, 2018: 499.
- luffae. Neoclystopsenella luffae Kurian, 1955: 133, figs. 249-255 (m.) INDIA (National Capital Territory).
- Combination in Tapinoma: Brown, 1988a: 337.
- Status as species: Shattuck, 1994: 146; Bolton, 1995b: 400.
- Junior synonym of melanocephalum: Guerrero, 2018: 500.
- malesiana. Tapinoma melanocephalum var. malesiana Forel, 1913k: 93 (w.q.) INDONESIA (Sumatra, Java).
- Subspecies of melanocephalum: Chapman & Capco, 1951: 192; Shattuck, 1994: 148; Bolton, 1995b: 400.
- Junior synonym of melanocephalum: Guerrero, 2018: 500.
- nana. Formica nana Jerdon, 1851: 125 (w.) INDIA (Karnataka; “found in all parts of India”).
- [Duplicated in Jerdon, 1854b: 108.]
- [Unresolved junior primary homonym of Formica nana Latreille, 1802c: 263 (Emery, 1921a: 26; Bolton, 1995b: 401).]
- Status as species: Smith, F. 1858b: 15; Mayr, 1863: 418; Smith, F. 1871a: 303.
- Junior synonym of melanocephalum: Emery, 1892b: 166; Forel, 1895e: 472.
- pellucida. Myrmica pellucida Smith, F. 1857a: 71 (w.) SINGAPORE.
- Status as species: Smith, F. 1858b: 124; Mayr, 1863: 434; Smith, F. 1871a: 324.
- Junior synonym of melanocephalum: Mayr, 1886c: 359; Forel, 1895e: 472.
Type Material
- Formica melanocephalum: Neotype (designated by Guerrero, 2018: 499), worker, Magdalena, Santa Marta, Calabazo, roadside to Pueblito-PNN Tayrona, Colombia, 11°16′57″N 74°01′26″W / 11.2825°N 74.0239°W, 22 January 2017, R. Guerrero, nest series, RJG01286 (ICNC-093716), Museo de Historia Natural; nest in dead branch on the roadside, secondary dry forest.
- Tapinoma australis: Syntype, 23 workers, Santo Island, Hog Harbour, Vanuatu, The Natural History Museum, Naturhistorisches Museum, Basel.
- Tapinoma melanocephalum coronatum: Lectotype (designated by Guerrero, 2018: 500), worker, Puntarenas, Costa Rica, CASENT0909775, Musee d'Histoire Naturelle Genève; middle specimen on the pin.
- Tapinoma melanocephalum coronatum: Paralectotype (designated by Guerrero, 2018: 500), 2 workers, Puntarenas, Costa Rica, Musee d'Histoire Naturelle Genève; upper and lower specimen on the same pin as the lectotype.
- Tapinoma melanocephalum malesiana: Lectotype (designated by Guerrero, 2018: 500), worker, Sumatra, Soengei Bamban, Indonesia, CASENT0909776, Musee d'Histoire Naturelle Genève.
- Tapinoma melanocephalum malesiana: Paralectotype (designated by Guerrero, 2018: 500), 45 workers, 6 queens, Sumatra, Soengei Bamban, Indonesia, CASENT0909776, Musee d'Histoire Naturelle Genève.
- Tapinoma melanocephalum malesiana: Paralectotype (designated by Guerrero, 2018: 500), 1 worker, Sumatra, Soengei Bamban, Indonesia, CASENT0909776, Naturhistorisches Museum, Basel.
- Neoclystopsenella luffae: Holotype, male, Delhi, India, 2 July 1938, Imperial Entomologist, Forest Research Institute, Dehradun, India; On Luffa; additional types: Paratypes: 2 males, same data of holotype; 1 male same data of the holotype, except on cowpea and 5-VII-1938 (see Singh et al., 2021).
The following notes on F. Smith type specimens have been provided by Barry Bolton (details):
Formica familiaris
Holotype queen in Oxford University Museum of Natural History. Labelled “Mak.” (= Makassar, Sulawesi).
Myrmica pellucida
Four worker syntypes in Oxford University Museum of Natural History. Without a locality label on the specimens, but with a Smith det. label. Locality given in original description is Singapore.
Description
Unless otherwise noted the text for the remainder of this section is reported from Guerrero, 2018.
Worker
Worker diagnosis. Bicolored small ants, with head and mesosoma black to brown, mandibles, antennae, legs and gaster light brown to pale yellow; third and fourth segment of the maxillary palps broad and flattened, spatulalike; basal end of the fifth segment inserted into the ventral face of the fourth segment.
Worker description (Figs 13–14): Head. Ovoid, longer than wide with slightly convex lateral margins; posterior margin variable, convex to straight, or sometimes slightly emarginated medially, in all cases, with rounded posterolateral corners. Mandibles subtriangular, straight outer margin over 2/3 length, and distally curved inwards; masticatory margin consisting of strong apical tooth, acute and longer than wide; slender subapical tooth, half length of the apical tooth; third triangular tooth half as long as subapical; fourth tooth slightly longer than third, followed by row of tight denticles decreasing in size approaching curvature leading to undifferentiated basal margin, the latter completely smooth or slightly crenulated. Palpal formula 6:4; maxillary palps with the first segment small and cylindrical, second segment conical and wider distally, sixth segment elongated, digitiform, rounded at tip; segments of labial palps relatively similar in size, 1st and 2nd segments wedge-shaped, 3rd subrectangular, last segment oval, longer than wide, with rounded tip. Clypeus broader than long; anteromedial margin with slight depression, and narrow, barely noticeable; margins lateral to depression relatively straight, those more lateral obliquely straight; posterior margin of clypeus convex, almost reaching base of frontal carinae. Frontal carinae diverging distally. Antennal scapes long, surpassing posterior margin of head by distance similar to length of pedicel, outer margins straight, slightly curved at both base and apex, wider near apical end; pedicel slightly conical with sub-parallel sides, third segment twice as wide as long, fourth-eleventh segments rectangular, much longer than wide, gradually elongating towards apex, last segment twice as long as others, rounded at tip. Eyes relatively large, oval, longer than wide, external margin relatively straight while the internal margin strongly convex, distance from the mandibular insertion approximately 1/2 maximum length of the eye.
Mesosoma. In lateral view, promesonotum strongly convex, forming nearly continuous line; metanotal sulcus broad and relatively deep; dorsopropodeum strongly convex, dome-shaped, at same level or slightly below level of promesonotum, declivitous face straight, inclined between 50–60º. Propodeal spiracle nearly circular, slightly longer than wide, opening laterally, well below half-length of lateropropodeum.
Metasoma. In lateral view, petiole short, flattened dorsal face with sharp apex inclined anteriorly; ventral lobe projecting, with straight anterior face and slightly curved posterior face. First tergum of gaster excavated anteriorly, as in other Tapinoma species. Body surface and pilosity. Cephalic surface opaque, finely covered with punctures and reticulations forming very small alveoli; abundant pubescence very short. Mandibles relatively smooth and slightly opaque, dorsal surface tenuously foveolate, covered with abundant short suberect hairs, and longer suberect hairs towards middle and basal portion; ventral face with long erect hairs. Dorsal face of clypeus covered with pubescence similar to that of cephalic capsule; anterior clypeal margin with two longer and thicker hairs, one on either side of medial depression, two golden relatively shorter setae than previously described hairs, above the clypeal margin are a pair of thick hairs directed anteriorly. Mesosoma completely covered with fine reticulations forming alveoli, relatively larger than those of head; surface rough and opaque, with dense whitish appressed hairs, very short and separated, sometimes imperceptible, giving appearance of glabrous surface, only visible in oblique-dorsal position. Legs with relatively fine granular and reticulated sculpture, relatively opaque, completely covered with dense whitish pubescence.
Petiole smooth and shiny, with few short, decumbent hairs near posteroventral margin. Gaster smooth to finely reticulated, relatively shiny, covered with fine dense golden yellowish pubescence; 5–6 erect hairs near posterior margin of fourth tergum, other terga without erect hairs; sternites with two erect hairs per segment. Coloration. Bicolored, variable in color tone. Head and mesosoma intense dark brown to brown with some regions yellowish brown on the propleura and the dorsum of the promesonotum. Mandibles, maxillary and labial palps, scapes, antennal segments, and legs pale yellow; masticatory margin reddish brown. Gaster brown to pale yellow as legs, but sometimes with contrasting brown bands on posterior margins of each tergum.
Neotype measurement: HL: 0.51 mm, HW: 0.45 mm, SL: 0.47 mm; WL: 0.44 mm; CI: 88; SI: 92.
Worker measurements (n= 316): HL: 0.46 (0.38–0.60) mm, HW: 0.41 (0.35–0.60) mm, SL: 0.43 (0.35–0.52) mm; WL: 0.39 (0.28–0.60) mm; CI: 89 (80–104); SI: 94 (82–105).
Gyne
Gyne measurements (n= 20): HL: 0.52 (0.43–0.60) mm; HW: 0.51 (0.41–0.60) mm; SL: 0.45 (0.35–0.52) mm; WL: 0.74 (0.59–0.83) mm; CI: 93–104; SI: 81–93.
Gyne diagnosis: Cephalic capsule diverging posteriorly, much wider posterior to the compound eyes than anterior to them. Second segment of maxillary palps triangular, flattened, longer than wide, 3rd and 5th segments much broader and flattened, 3rd and 4th segment almost fused at the ends where they meet, the basal end of the 5th segment inserted into the ventral face of the 4th segment.
Gyne description (Figs 15–16): Head. Capsule cephalic square to rectangular (CI: 93–104), cuneiform; sides of the capsule relatively straight, lateral margin interrupted by the outer margin of the eyes; well-defined posterolateral corners, rounded, continuing to straight posterior cephalic margin, with shallow and wide concavity. Mandibles as in workers. Palpal formula 6:4; first segment of maxillary palps cylindrical and thickened, last segment thickened, digitiform, rounded at tip; segments of labial palps as in workers, but relatively broader and flattened. Dorsal face of clypeus as in workers, except for anterior margin relatively straighter and entire, with a slight mesal concavity. Frontal carinae divergent posteriorly. Scapes long, surpassing posterolateral corners of head by distance similar to length of pedicel; outer and inner margins straight in most of the length but slightly curved distally, wider mid-length; antennal segments as in workers. Compound eyes large, located at middle of length of cephalic capsule, relatively close to clypeal insertion; eyes located laterally, external margin breaking border of the cephalic capsule. Ocelli small, near level of posterior border of head; posterior ocelli near vertex but never breaking vertexal line.
Mesosoma. In profile, pronotum broad, propleura relatively broader than anterior part of pronotum; dorsal surface of mesonotum flattened; metanotum consisting of very thin strip, separated by deep mesonotal groove; dorsopropodeum without distinctive face, bending abruptly on a relatively inclined slope; mesopleuron divided into anepisternum and katepisternum, those divided by marked and relatively deep anapleural sulcus, anepisternum inverted U-shape, katepisternum subquadrate, almost as long as wide; propodeal spiracle circular, with an outer rim like a ring, directed posterad, and located on lateropropodeal face; metapleuron protruding, forming slight depression below propodeal spiracle. In dorsal view, mesoscutum strongly convex anteriorly, with parallel sides, diverging slightly posteriorly; mesonotal groove straight, slightly marked; mesoscutellum wider than long, posterior margin convex; dorsopropodeum restricted to very narrow band, posterior margin rounded bended abruptly on a slope, propodeum with the lateral margins, almost parallel.
Metasoma. Petiole relatively large, longer than wide, petiolar scale very low, restricted to small dorsal projection, inclined, directed anteriorly, with poorly developed lateral carinae; developed ventral lobe, most prominent posteriorly. Gaster as long as the mesosoma, flattened dorsally and slightly convex on ventral face. Body surface and pilosity. Surface of the head dense and finely punctate; with dense whitish decumbent hairs relatively separate, distance between hairs less than individual length. Dorsal face of mandibles densely covered with short golden hairs, on almost entire surface, except near area behind masticatory margin where they are long and inwardly directed. Maxillary palps covered with very short pilosity; labial palps with short erect and suberect hairs. Clypeus with pubescence as remainder of head, pattern of hairs on anterior margin as described in workers. Mesosoma opaque, abundantly and finely punctate; lateral regions of mesosoma covered with whitish pubescence, relatively longer than those of head, more abundant on katepisternum and the metapleuron; dorsum of the mesosoma covered with short pubescence, widely spaced, leaving surface partially exposed. Legs covered with short and decumbent hairs. Metasomal surface finely punctate as mesosoma, but relatively brighter; covered with abundant pubescence; fourth tergite with six long erect hairs near posterior margin, one central pair, and other two towards sides of the tergite.
Color. Head brown to dark brown, mandibles, maxillary and labial palps and antennae pale yellow; mesosoma bicolored, part of mesoscutum, mesoscutellum and legs yellowish brown to pale yellow, dorsum of pronotum and central portion of mesoscutum as spot, mesopleuron, inferior part of propodeum, and metapleuron brown. Petiole yellowish; gaster brown with yellowish brown bands on posterior margin
Male
Male measurements (n= 17): HL: 0.44 (0.43–0.46) mm; HW: 0.42 (0.41–0.45) mm; SL: 0.39 (0.35–0.43) mm; WL: 0.66 (0.59–0.73) mm; CI: 93–104; SI: 81–93.
Male diagnosis: Third and fourth segment of the maxillary palps widened, oblong in shape, much longer than wide; the basal end of the 5th segment inserted into the ventral face of the 4th segment, like in the female caste.
Male description (Figs 17–18): Head. In dorsal view, head as long as broad, largest width posterior to compound eyes, with lateral margin convex, cephalic capsule converging anteriorly, posterolateral corners rounded, continuous with straight posterior cephalic margin; ventral surface of cephalic capsule relatively flattened. Mandibles long, semifalcate; external margin slightly sinuous, curving inwards distally; masticatory margin straight, with apical tooth relatively large, obliquely directed downwards, subapical denticle overhanging from margin, remainder of margin with very tight denticles, same size, continuing towards the undifferentiated basal margin. Palpal formula 6:4; 1st segment cylindrical, 2nd segment two times longer than first, cuneiform, 4th segment much wider than 3rd, 5th and 6th segments finger-shaped, the apical with rounded tip. Clypeus broad, relatively high, dorsal face outwardly projected; anteromedial border slightly concave, narrow, with rounded corners continuing toward relatively straight anterior margin, anterolateral margins inclined posteriorly; posterior margin convex but posterior vertex relatively flattened. Frontal carinae straight and semi-parallel at base, curved and diverging distally. Scape long, surpassing posterior cephalic margin by distance slightly less than distance between frontal carinae, scape with lateral margins parallel, outwardly curved from base to medial length, straight for rest of length, pedicel cuneiform, narrower at base, remainder of antennal segments cylindrical, longer than broad, last segment almost twice length of others. Compound eyes very large, oval, two times longer than broad, occupying almost entire lateral portion of head, breaking outline of lateral margins; anterior margin of eye separated from clypeal insertion by distance shorter than minimal width of scape. Ocelli large, located on wide base and relatively higher than dorsal surface of head, slightly below posterior cephalic vertex; anteromedial ocellus relatively smaller than lateral ocelli.
Mesosoma. In profile, propleura broad, flattened, with dorsal border thin, projecting dorsally; mesonotum anteriorly curved, with hump in lateral view, dorsal face relatively straight and high; mesopleuron divided into anepisternum and katepisternum, both delimited by wide and relatively deep anapleural sulcus; anepisternum inverted U-shaped, relatively wider than long, with dorsal margin slightly inclined posteriorly, anterior margin longer than posterior; katepisternum semiquadrate. Dorsopropodeum below level of mesonotum, with long surface, posteropropodeum slightly bulging decline of 90° toward base; propodeal spiracle oval, longer than broad, located on lateropropodeum, opening posteriorly. In dorsal view, anterior line of pronotum visible; mesoscutum slightly triangular with inferior margin truncated, longer than broad; dorsal face of propodeum visible.
Metasoma. In lateral view, petiole with petiolar scale well developed, anterior face relatively high and straight, forming angle with oblique dorsal face that descends posteriorly to declivity; ventral lobe developed, slightly convex. Cupula weakly sclerotized, easily collapsible when specimen pinned; telomeres dorsoventrally broad, obliquely truncated at apex; digitus long and narrow to tip, latter ventrally curved, cuspis located well inside genital capsule, semi-rectangular in lateral/medial view. Body surface and pilosity. Head surface opaque, softly punctulate; abundant, whitish, appressed hairs. Mandibles smooth, shining, bearing scattered short and long hairs, subdecumbent from base to behind masticatory margin; some hairs long, surpassing external margin. Anterior margin of clypeus with same pattern of hairs as described in worker; more lateral portions with relatively larger curved hairs; dorsal face densely hairy. Scape surface opaque, slightly punctate, covered by very short pilosity, with some hairs relatively longer and suberect in lateral margins; antennal segments with surface granulate, densely covered by short decumbent hairs, with some short suberect hairs.
Mesosoma relatively shining, with some areas notably opaque, puncticulate; surface covered by relatively short dense golden scattered pubescence, more abundant on katepisternum and metapleuron. Gaster reticulate, with small alveoli; covered by dense golden pubescence, but relatively sparse, leaving uncovered surface; telomeres densely covered by relatively long pilosity, surpassing margin.
Color. Ants completely yellowish brown, except for mandibles, scape and antennal segment, and legs which are whitish, contrasting with the remainder of body.
Larvae: Dolichoderine larvae described by Wheeler and Wheeler (1951), and examined in detail by Jesus et al. (2010).
Karyotype
- See additional details at the Ant Chromosome Database.
- Explore: Show all Karyotype data or Search these data. See also a list of all data tables or learn how data is managed.
- 2n = 10, karyotype = 8M+2A (India) (Imai et al., 1984).
- 2n = 10 (Indonesia) (Imai et al., 1985).
- n = 5, 2n = 10 (USA) (Crozier, 1970a; Crozier, 1970b).
References
- Abdar, M.R. 2020. Seasonal abundance and commonly occurring household ants species in Sangli District Maharashtra. Research Journal of Agricultural Sciences 11(6): 1413-1415.
- Abdel-Dayem, M., Al Dhafer, H., Aldawood, A., Sharaf, M. 2021. An update to the taxonomy and distribution of the Arabian Tapinoma Foerster, 1850 (Hymenoptera: Formicidae) with an illustrated key and remarks on habitats. Biodiversity Data Journal 9, e66058 (doi:10.3897/bdj.9.e66058).
- Ahmad, Z., Ghramh, H.A., Ali Khan, K., Khan, F., Shujauddin. 2020. Impact of Two Attending Ants, Crematogaster subnuda and Camponotus compressus (Hymenoptera: Formicidae), on the Parasitism of Sugarcane Aphid Melanaphis sacchari (Zehnt.) Pakistan Journal of Zoology (doi:10.17582/journal.pjz20200309190337).
- Akhila, A., Keshamma, E. 2022. A review on diversity of ant species in Karnataka State. International Journal of Advanced Scientific Research and Management 7(6): 15-20 (doi:10.36282/IJASRM/7.6.2022.1865).
- Albuquerque, E., Prado, L., Andrade-Silva, J., Siqueira, E., Sampaio, K., Alves, D., Brandão, C., Andrade, P., Feitosa, R., Koch, E., Delabie, J., Fernandes, I., Baccaro, F., Souza, J., Almeida, R., Silva, R. 2021. Ants of the State of Pará, Brazil: a historical and comprehensive dataset of a key biodiversity hotspot in the Amazon Basin. Zootaxa 5001, 1–83 (doi:10.11646/zootaxa.5001.1.1).
- Baidya, P., Bagchi, S. 2021. Influence of human land use and invasive species on beta diversity of tropical ant assemblages. Insect Conservation and Diversity, icad.12536 (doi:10.1111/icad.12536).
- Baltazar, C.R. 1966. A catalogue of Philippine Hymenoptera (with a bibliography, 1758-1963). Pacific Insects Monographs 8: 1-488. (page 263, listed)
- Báthori, F., Herczeg, G., Vilizzi, L., Jégh, T., Kakas, C., Petrovics, M., Csősz, S. 2024. A survey and risk screening of non-native ant species colonising greenhouses in Hungary. Biological Invasions (doi:10.1007/s10530-023-03227-9).
- Baty, J.W., Bulgarella, M., Dobelmann, J., Felden, A., Lester, P.J. 2020. Viruses and their effects in ants (Hymenoptera: Formicidae). Myrmecological News 30: 213-228 (doi:10.25849/MYRMECOL.NEWS_030:213).
- Berville, L., Hefetz, A., Espadaler, X., Lenior, A., Renuuci, M., Blight, O., Provost, E. 2013. Differentiation of the ant genus Tapinoma (Hymenoptera: Formicidae) from the Mediterranean Basin by species-specific cuticular hydrocarbon profiles. Myrmecological News 18: 77-92.
- Bharti, H., Sharma, Y.P., Kaur, A. 2009. Seasonal patterns of ants (Hymenoptera: Formicidae) in Punjab Shivalik. Halteres 1: 36-47.
- Blard, F., Dorow, W.-H.-O., Delabie, J. H. C. 2003. Les Fourmis de l’île de la Réunion (Hymenoptera, Formicidae). Bulletin de La Société Entomologique de France, 108(2), 127–137 (doi:10.3406/bsef.2003.16939).
- Bolton, B. 1995b. A new general catalogue of the ants of the world. Cambridge, Mass.: Harvard University Press, 504 pp. (page 401, Senior synonym of australe: new synonymy)
- Borowiec, L. 2014. Catalogue of ants of Europe, the Mediterranean Basin and adjacent regions (Hymenoptera: Formicidae). Genus (Wroclaw) 25(1-2): 1-340.
- Borowiec, L., Salata, S. 2018. Notes on ants (Hymenoptera: Formicidae) from Gambia (Western Africa). Annals of the Upper Silesian Museum in Bytom, Entomology 26 (online 010), 1-13 (doi:10.5281/ZENODO.1243767).
- Borowiec, L., Salata, S. 2019. Next step in the invasion: Trichomyrmex mayri (Forel, 1902) new to the Philippines (Hymenoptera: Formicidae). Annals of the Upper Silesian Museum in Bytom Entomology 28:1-3 (doi:10.5281/ZENODO.2644912).
- Brassard, F., Leong, C.-M., Chan, H.-H., Guénard, B. 2021. High diversity in urban areas: How comprehensive sampling reveals high ant species richness within one of the most urbanized regions of the world. Diversity 13, 358 (doi:10.3390/d13080358).
- Cantone S. 2017. Winged Ants, The Male, Dichotomous key to genera of winged male ants in the World, Behavioral ecology of mating flight (self-published).
- Cantone S. 2018. Winged Ants, The queen. Dichotomous key to genera of winged female ants in the World. The Wings of Ants: morphological and systematic relationships (self-published).
- Cheng, D. F., S. Q. Chen, Y. Q. Huang, N. E. Pierce, M. Riegler, F. Yang, L. Zeng, Y. Y. Lu, G. W. Liang, and Y. J. Xu. 2019. Symbiotic microbiota may reflect host adaptation by resident to invasive ant species. Plos Pathogens. 15:22. doi:10.1371/journal.ppat.1007942
- Collingwood, C. A. 1979. The Formicidae (Hymenoptera) of Fennoscandia and Denmark. Fauna Entomol. Scand. 8:1-174.
- Collingwood, C.A., Agosti, D., Sharaf, M.R., van Harten, A. 2011. Order Hymenoptera, family Formicidae. Arthropod fauna of the UAE 4: 405-474.
- Crozier, R. H. 1970a. Karyotypes of twenty-one ant species (Hymenoptera: Formicidae), with reviews of the known ant karyotypes. Can. J. Genet. Cytol. 12: 109-128 (page 119, karyotype described)
- Csősz, S., Báthori, F., Gallé, L., Lőrinczi, G., Maák, I., Tartally, A., Kovács, É., Somogyi, A.Á., Markó, B. 2021. The myrmecofauna (Hymenoptera: Formicidae) of Hungary: Survey of ant species with an annotated synonymic inventory. Insects 16;12(1):78 (doi:10.3390/insects12010078).
- Csosz, S., Marko, B., Galle, L. 2011. The myrmecofauna (Hymenoptera: Formicidae) of Hungary: an updated checklist. North-Western Journal of Zoology 7: 55-62.
- Debout, G., Schatz, B., Elias, M., Mckey, D. 2007. Polydomy in ants: What we know, what we think we know, and what remains to be done. Biological Journal of the Linnean Society 90: 319-348.
- Dekoninck, W., Ignace, D., Vankerkhoven, F., Wegnez, P. 2012. Verspreidingsatlas van de mieren van België. Bulletin de la Société royale belge d’Entomologie 148: 95-186.
- Dekoninck, W., Wauters, N., Delsinne, T. 2019. Capitulo 35. Hormigas invasoras en Colombia. Hormigas de Colombia.
- Deyrup, M., Davis, L. & Cover, S. 2000. Exotic ants in Florida. Transactions of the American Entomological Society 126, 293-325.
- Deyrup, M.A., Carlin, N., Trager, J., Umphrey, G. 1988. A review of the ants of the Florida Keys. Florida Entomologist 71: 163-176.
- Dias, R.K.S., Kosgamage, K.R.K.A. 2013. Occurrence and species diversity of ground-dwelling worker ants (Family: Formicidae) in selected lands in the dry zone of Sri Lanka. Journal of Science of the University of Kelaniya Sri Lanka 7: 55-72 (doi:10.4038/josuk.v7i0.6233).
- Dias, R.K.S., Rajapaksa, R.P.K.C. 2017. Geographic records of subfamilies, genera and species of ants (Hymenoptera: Formicidae) in the four climatic zones of Sri Lanka: A review. Journal of Science of the University of Kelaniya Sri Lanka 11, 23-45. (doi:10.4038/josuk.v11i2.7999).
- do Nascimento, L.E., Amaral, R.R., Ferreira, R.M.dos A., Trindade, D.V.S., do Nascimento, R.E., da Costa, T.S., Souto, R.N.P. 2020. Ants (Hymenoptera: Formicidae) as potential mechanical vectors of pathogenic bacteria in a public hospital in the Eastern Amazon, Brazil. Journal of Medical Entomology 57: 1619–1626. (doi:10.1093/JME/TJAA062).
- Dubovikoff, D.A., Yusupov, Z.M. 2017. Family Formicidae - Ants. In Belokobylskij S. A. and A. S. Lelej: Annotated catalogue of the Hymenoptera of Russia. Proceedingss of the Zoological Institute of the Russian Academy of Sciences 6: 197-210.
- Ellison, A.M., Gotelli, N.J. 2021. Ants (Hymenoptera: Formicidae) and humans: from inspiration and metaphor to 21st-century symbiont. Myrmecological News 31: 225-240 (doi:10.25849/MYRMECOL.NEWS_031:225).
- Emery, C. 1887a [1886]. Catalogo delle formiche esistenti nelle collezioni del Museo Civico di Genova. Parte terza. Formiche della regione Indo-Malese e dell'Australia. [part]. Ann. Mus. Civ. Stor. Nat. 24[=(2)(4): 209-240 (page 249, male described)
- Emery, C. 1892c [1891]. Note sinonimiche sulle formiche. Bull. Soc. Entomol. Ital. 23: 159-167 (page 166, Senior synonym of nana)
- Espadaler, X. 2007. The ants of El Hierro (Canary Islands). Pages 113-127 in R. R. Snelling, B. L. Fisher, and P. S. Ward, editors. Advances in ant systematics (Hymenoptera: Formicidae): homage to E. O. Wilson - 50 years of contributions. Memoirs of the American Entomological Institute, Gainesville, FL. 80:690 pp.
- Eyer, P.A., Vargo, E.L. 2021. Breeding structure and invasiveness in social insects. Current Opinion in Insect Science 46: 24-30.
- Fabricius, J. C. 1793. Entomologia systematica emendata et aucta. Secundum classes, ordines, genera, species, adjectis synonimis, locis observationibus, descriptionibus. Tome 2. Hafniae [= Copenhagen]: C. G. Proft, 519 pp. (page 353, worker described)
- Fontenla, J.L., Brito, Y.M. 2011. Hormigas invasoras y vagabundas de Cuba. Fitosanidad 15(4), 253-259.
- Forel, A. 1891c. Les Formicides. [part]. In: Grandidier, A. Histoire physique, naturelle, et politique de Madagascar. Volume XX. Histoire naturelle des Hyménoptères. Deuxième partie (28e fascicule). Paris: Hachette et Cie, v + 237 pp. (page 102, queen described)
- Forel, A. 1899f. Formicidae. [part]. Biol. Cent.-Am. Hym. 3: 81-104 (page 101, Senior synonym of familaris)
- Franco, W., Ladino, N., Delabie, J.H.C., Dejean, A., Orivel, J., Fichaux, M., Groc, S., Leponce, M., Feitosa, R.M. 2019. First checklist of the ants (Hymenoptera: Formicidae) of French Guiana. Zootaxa 4674, 509–543 (doi:10.11646/zootaxa.4674.5.2).
- García-Avendaño, E.I. & Guerrero, R.J. 2018. Taxonomía y distribución de las hormigas del género Tapinoma (Formicidae: Dolichoderinae) en Colombia. Revista Colombiana de Entomología 44 (2): 223-237 (doi:10.25100/socolen.v44i2.7324).
- Guerrero, R.J. 2018. Taxonomic identity of the ghost ant, Tapinoma melanocephalum (Fabricius, 1793) (Formicidae: Dolichoderinae). Zootaxa 4410 (3): 497–510 (DOI 10.11646/zootaxa.4410.3.4).
- Guerrero, R.J. 2021. Transfer of two South American ant species from Tapinoma Foerster 1850 to Forelius Emery 1888 (Hymenoptera: Formicidae: Dolichoderinae). Zootaxa 4920, 428–438 (doi:10.11646/zootaxa.4920.3.8).
- Gull-E-Fareen, A., Bodlah, I., Rasheed, M. T., Niaz, Y., Bodlah, M. A., Asif, M., Khokhar, N. M. (2020). Trophic associations of ants with aphid partners and new distribution records of some ants in Pothwar Region of Pakistan. Pakistan Journal of Zoology, 531 (doi:10.17582/journal.pjz/20190510120507).
- Hammer, S., Jensen, J.-K. 2021. Discoveries and fate of six ant (Hymenoptera, Formicidae) species on the Faroe Islands. BioInvasions Records 10, 28–32 (doi:10.3391/bir.2021.10.1.04).
- Hasin, S., Tasen, W. 2020. Ant community composition in urban areas of Bangkok, Thailand. Agriculture and Natural Resources 54: 507-514 (doi:10.34044/j.anres.2020.54.5.07).
- Herrera, H.W., Baert, L., Dekoninck, W., Causton, C.E., Sevilla, C.R., Pozo, P., Hendrickx, F. 2020. Distribution and habitat preferences of Galápagos ants (Hymenoptera: Formicidae). Belgian Journal of Entomology, 93: 1–60.
- Herwina, H., Mairawita, Yulvita, L., Putri, D., Satria, R., Janra, M.N., Yaherwandi, Sakamaki, Y. 2020. Ant species composition (Hymenoptera: Formicidae) at a Highland Agricultural Area for Wheat and Potato in Alahan Panjang, West Sumatera. IOP Conference Series: Earth and Environmental Science 515, 012018 (doi:10.1088/1755-1315/515/1/012018).
- Heterick, B.E. 2021. A guide to the ants of Western Australia. Part I: Systematics. Records of the Western Australian Museum, Supplement 86, 1-245 (doi:10.18195/issn.0313-122x.86.2021.001-245).
- Heterick, B.E. 2022. A guide to the ants of Western Australia. Part II: Distribution and biology. Records of the Western Australian Museum, supplement 86: 247-510 (doi:10.18195/issn.0313-122x.86.2022.247-510).
- Hisasue, Y., Tsuji, Y. 2020. Records of Nylanderia amia (Forel) in Shikoku, with notes of range expansion of Japan. ARI 41: 18-36.
- Hoffmann, B., Eldridge, J., Marston, C. 2023. The first eradication of an exotic ant species from the entirety of Australia: Pheidole fervens. Management of Biological Invasions, 14(4), 619–624 (doi:10.3391/mbi.2023.14.4.03).
- Hosoishi, S., Rahman, M. M., Heng, S. 2022. Exotic ants (Hymenoptera: Formicidae) of Cambodia. Far Eastern Entomologist 460, 15–24 (doi:10.25221/fee.460.3).
- Imai, H.T., Kihara, A., Kondoh, M., Kubota, M., Kuribayashi, S., Ogata, K., Onoyama, K., Taylor, R.W., Terayama, M., Yoshimura, M., Ugawa, Y. 2003. Ants of Japan. 224 pp, Gakken, Japan.
- Ivanov, K. 2019. The ants of Ohio (Hymenoptera, Formicidae): an updated checklist. Journal of Hymenoptera Research 70: 65–87 (doi:10.3897@jhr.70.35207).
- Iwata, K., Eguchi, K., Yamane, S. 2005. A case study on urban ant fauna of southern Kyusyu, Japan, with notes on a new monitoring protocol (Insecta, Hymenoptera, Formicidae). Journal of Asia-Pacific Entomology 8, 263-272.
- Karaman, C., Kıran, K. 2018. New tramp ant species for Turkey: Tetramorium lanuginosum Mayr (Hymenoptera: Formicidae). Trakya University Journal of Natural Sciences 19(1), e1-e4 (doi:10.23902/trkjnat.340008).
- Katayama, M., Tsuji, K. 2010. Habitat differences and occurrence of native and exotic ants on Okinawa Island. Entomological Science 13, 425–429 (doi:10.1111/j.1479-8298.2010.00400.x).
- Khachonpisitsak, S., Yamane, S., Sriwichai, P., Jaitrong, W. 2020. An updated checklist of the ants of Thailand (Hymenoptera, Formicidae). ZooKeys 998, 1–182 (doi:10.3897/zookeys.998.54902).
- Khazan, E., Bujan, J., Scheffers, B.R. 2020. Patterns of ant activity and nesting ecology depend on flooding intensity in a Neotropical floodplain. International Journal of Tropical Insect Science (doi:10.1007/s42690-020-00149-0).
- Klimes, P. and J. Okrouhlik. 2015. Invasive ant Tapinoma melanocephalum (Hymenoptera: Formicidae): A rare guest or increasingly common indoor pest in Europe? European Journal of Entomology. 112:705-712. doi:10.14411/eje.2015.089
- Kreider, J.J., Chen, T.W., Hartke, T.R., Buchori, D., Hidayat, P., Nazarreta, R., Scheu, S., Drescher, J. 2021. Rainforest conversion to monocultures favors generalist ants with large colonies. Ecosphere 12 (doi:10.1002/ecs2.3717).
- Latumahina, F., Borovanska, M., Musyafa, Sumardi, Susetya Putra, N., Janda, M. 2015. Ants of Ambon Island – diversity survey and checklist. ZooKeys 472, 43–57 (doi:10.3897/zookeys.472.8441).
- Lee, C.-C., Weng, Y.-M., Lai, L.-C., Suarez, A.V., Wu, W.-J., Lin, C.-C., Yang, C.-C.S. 2020. Analysis of recent interception records reveals frequent transport of arboreal ants and potential predictors for ant invasion in Taiwan. Insects 11, 356 (doi:10.3390/INSECTS11060356).
- Liu, C., Fischer, G., Hita Garcia, F., Yamane, S., Liu, Q., Peng, Y.Q., Economo, E.P., Guénard, B., Pierce, N.E. 2020. Ants of the Hengduan Mountains: a new altitudinal survey and updated checklist for Yunnan Province highlight an understudied insect biodiversity hotspot. ZooKeys 978, 1–171 (doi:10.3897/zookeys.978.55767).
- Lubertazzi, D. 2019. The ants of Hispaniola. Bulletin of the Museum of Comparative Zoology, 162(2), 59-210 (doi:10.3099/mcz-43.1).
- MacGown, J.A. 2017. First record of the Ghost Ant, Tapinoma melanocephalum (Hymenoptera: Formicidae), from Alabama. Transactions American Entomological Society 143: 533-535.
- MacGown, J.A., Booher, D., Richter, H., Wetterer, J.K., Hill, J.G. 2021. An updated list of ants of Alabama (Hymenoptera: Formicidae) with new state records. Transactions of the American Entomological Society 147: 961-981 (doi:10.3157/061.147.0409).
- Mayr, G. 1862. Myrmecologische Studien. Verh. K-K. Zool.-Bot. Ges. Wien 12: 649-776 (page 651, Combination in Tapinoma)
- Mayr, G. 1886c. Notizen über die Formiciden-Sammlung des British Museum in London. Verh. K-K. Zool.-Bot. Ges. Wien 36: 353-368 (page 359, Senior synonym of pellucida)
- Melo, T.S., Koch, E.B.A., Andrade, A.R.S., Travassos, M.L.O., Peres, M.C.L., Delabie, J.H.C. 2021. Ants (Hymenoptera: Formicidae) in different green areas in the metropolitan region of Salvador, Bahia state, Brazil. Brazilian Journal of Biology 82, e236269 (doi:10.1590/1519-6984.236269).
- Meurgey, F. 2020. Challenging the Wallacean shortfall: A total assessment of insect diversity on Guadeloupe (French West Indies), a checklist and bibliography. Insecta Mundi 786: 1–183.
- Molfini, M., Zapparoli, M., Genovesi, P., Carnevali, L., Audisio, P., Di Giulio, A., Bologna, M.A. 2020. A preliminary prioritized list of Italian alien terrestrial invertebrate species. Biological Invasions 22(8), 2385–2399 (doi:10.1007/s10530-020-02274-w).
- Moura, M.N., Cardoso, D.C., Cristiano, M.P. 2020. The tight genome size of ants: diversity and evolution under ancestral state reconstruction and base composition. Zoological Journal of the Linnean Society, zlaa135 (doi:10.1093/zoolinnean/zlaa135).
- Mukherjee, A., Roy, U.S. 2021. Role of physicochemical parameters on diversity and abundance of ground–dwelling, diurnal ant species of Durgapur Government College Campus, West Bengal, India. Munis Entomology and Zoology Journal 16(1): 366-380.
- Nalini, T., Jayanthi, R. 2020. First record on interaction between ants, other insect visitors and extrafioral nectaries (EFNS) in selected crops from India. Plant Archives 20, 2105-2113.
- Narváez-Vásquez, A., Gaviria, J., Vergara-Navarro, E.V., Rivera-Pedroza, L., Löhr, B. 2021. Ant (Hymenoptera: Formicidae) species diversity in secondary forest and three agricultural land uses of the Colombian Pacific Coast. Revista Chilena de Entomologia 47, 441–458 (doi:10.35249/rche.47.3.21.01).
- Nazarreta, R., Hartke, T.R., Hidayat, P., Scheu, S., Buchori, D., Drescher, J. 2020. Rainforest conversion to smallholder plantations of rubber or oil palm leads to species loss and community shifts in canopy ants (Hymenoptera: Formicidae). Myrmecological news 30, 175-186 (doi:10.25849/MYRMECOL.NEWS_030:175).
- Nooten, S.S., Lee, R.H., Guénard, B. 2021. Evaluating the conservation value of sacred forests for ant taxonomic, functional and phylogenetic diversity in highly degraded landscapes. Biological Conservation 261, 109286 (doi:10.1016/j.biocon.2021.109286).
- Obregon, R., M. R. Shaw, J. Fernandez-Haeger, and D. Jordano. 2015. Parasitoid and ant interactions of some Iberian butterflies (Insecta: Lepidoptera). Shilap-Revista De Lepidopterologia. 43:439-454.
- Oussalah, N., Marniche, F., Espadaler, X., Biche, M. 2019. Exotic ants from the Maghreb (Hymenoptera, Formicidae) with first report of the hairy alien ant Nylanderia jaegerskioeldi (Mayr) in Algeria. Arxius de Miscel·lània Zoològica, 45–58 (doi:10.32800/amz.2019.17.0045).
- Rafael, J.A., Limeira-de-Oliveira, F., Hutchings, R.W., Miranda, G.F.G., Silva Neto, A.M.da, Somavilla, A., Camargo, A., Asenjo, A., Pinto, Â.P., Bello, A.de M., Dalmorra, C., Mello-Patiu, C.A.de, Carvalho, C.J.B.de, Takiya, D.M., Parizotto, D.R., Marques, D.W.A., Cavalheiro, D.de O., Mendes, D.M.de M., Zeppelini, D., Carneiro, E., Lima, É.F.B., Lima, E.C.A.de, Godoi, F.S.P.de, Pessoa, F.A.C., Vaz-de-Mello, F.Z., Sosa-Duque, F.J., Flores, H.F., Fernandes, I.O., Silva-Júnior, J.O., Gomes, L.R.P., Monné, M.L., Castro, M.C.M.de, Silva, M.P.G.da, Couri, M.S., Gottschalk, M.S., Soares, M.M.M., Monné, M.A., Rafael, M.S., Casagrande, M.M., Mielke, O.H.H., Grossi, P.C., Pinto, P.J.C., Bartholomay, P.R., Sobral, R., Heleodoro, R.A., Machado, R.J.P., Corrêa, R.C., Hutchings, R.S.G., Ale-Rocha, R., Santos, S.D.dos, Lima, S.P.de, Mahlmann, T., Silva, V.C., Fernandes, D.R.R. 2020. Insect (Hexapoda) diversity in the oceanic archipelago of Fernando de Noronha, Brazil: updated taxonomic checklist and new records. Revista Brasileira de Entomologia 64, e20200052 (doi:10.1590/1806-9665-rbent-2020-0052).
- Ramalho, M.de O., Kim, Z., Wang, S., Moreau, C.S. 2021. Wolbachia Across Social Insects: Patterns and Implications. Annals of the Entomological Society of America 114, 206–218 (doi:10.1093/aesa/saaa053).
- Rasheed, M.T., Bodlah, I., Fareen, A.G., Wachkoo, A.A., Huang, X., Akbar, S.A. 2019. A checklist of ants (Hymenoptera: Formicidae) in Pakistan. Sociobiology 66(3), 426-439 (doi:10.13102/sociobiology.v66i3.4330).
- Reyes, J.L. 2010. Apuntes sobre una comunidad de hormigas sinantrópicas en Santiago de Cuba (Hymenoptera: Formicidae). Cocuyo 18: 44-47.
- Roger, J. 1862b. Beiträge zur Kenntniss der Ameisenfauna der Mittelmeerländer. II. Berl. Entomol. Z. 6: 255-262 (page 258, Combination in Micromyrma)
- Rosas-Mejía, M., Guénard, B., Aguilar-Méndez, M. J., Ghilardi, A., Vásquez-Bolaños, M., Economo, E. P., Janda, M. 2021. Alien ants (Hymenoptera: Formicidae) in Mexico: the first database of records. Biological Invasions 23(6), 1669–1680 (doi:10.1007/s10530-020-02423-1).
- Santschi, F. 1928e. Nouvelles fourmis d'Australie. Bull. Soc. Vaudoise Sci. Nat. 56: 465-483 (page 475, Combination in Tapinoma (Micromyrma))
- Seifert, B. 2022. The previous concept of the cosmopolitan pest ant Tapinoma melanocephalum (Fabricius, 1793) includes two species (Hymenoptera: Formicidae: Tapinoma). Osmia 10: 35-44 (doi:10.47446/OSMIA10.4).
- Seifert, B., Kaufmann, B., Fraysse, L. 2024. A taxonomic revision of the Palaearctic species of the ant genus Tapinoma Mayr 1861 (Hymenoptera: Formicidae). Zootaxa 5435(1), 1-74 (doi:10.11646/zootaxa.5435.1.1).
- Shakur, M.A., Mangaly, P., Baidya, P., Bagchi, S. 2024. A new species of the genus Tapinoma Foerster, 1850 (Hymenoptera: Formicidae) from the Western Ghats, India. Asian Myrmecology 17, e017005 (doi:10.20362/AM.017005).
- Sharaf, M. R., Wetterer, J. K., Mohamed, A. A., Aldawood, A. S. 2022. Faunal composition, diversity, and distribution of ants (Hymenoptera: Formicidae) of Dhofar Governorate, Oman, with updated list of the Omani species and remarks on zoogeography. European Journal of Taxonomy 838: 1-106 (doi:10.5852/ejt.2022.838.1925).
- Sharaf, M.R., Abdel-Dayem, M.S., Mohamed, A.A., Fisher, B.L., Aldawood, A.S. 2020. A preliminary synopsis of the ant fauna (Hymenoptera: Formicidae) of Qatar with remarks on the zoogeography. Annales Zoologici 70: 533-560 (doi:10.3161/00034541anz2020.70.4.005).
- Sharaf, M.R., Aldawood, A.S. 2022. Tapinoma yacoubi, sp. nov., a new ant species (Hymenoptera: Formicidae) from Saudi Arabia, Zoology in the Middle East, 68:2, 165-170 (doi:10.1080/09397140.2022.2077565).
- Sharaf, M.R., Fisher, B.L., Collingwood, C.A., Aldawood, A.S. 2017. Ant fauna (Hymenoptera: Formicidae) of the Socotra Archipelago (Yemen): zoogeography, distribution and description of a new species. Journal of Natural History 51, 317–378 (DOI 10.1080/00222933.2016.1271157).
- Shattuck, S. O. 1994. Taxonomic catalog of the ant subfamilies Aneuretinae and Dolichoderinae (Hymenoptera: Formicidae). Univ. Calif. Publ. Entomol. 112:i-xix, 1-241. (page 148, see also)
- Shi, Q. X., L. H. Lu, Y. Y. Lei, Y. R. He, and J. Chen. 2017. Gland Origin and Electroantennogram Activity of Volatile Compounds in Ghost Ants, Tapinoma melanocephalum (Hymenoptera: Formicidae) and Behavioral Response to (Z)-9-Nonadecene. Environmental Entomology. 46:1374-1380. doi:10.1093/ee/nvx164
- Shi, Q., He, Y., Chen, J., Lu, L. 2020. Thermally Induced Actinidine Production in Biological Samples. Journal of Agricultural and Food Chemistry 68, 12252–12258 (doi:10.1021/acs.jafc.0c02540).
- Siddiqui, J. A., Li, J., Zou, X., Bodlah, I., Huang, X. 2019. Meta-analysis of the global diversity and spatial patterns of aphid-ant mutualistic relationships. Applied Ecology and Environmental Research 17: 5471-5524 (doi:10.15666/aeer/1703_54715524).
- Siddiqui, J.A., Bamisile, B.S., Khan, M.M., Islam, W., Hafeez, M., Bodlah, I., Xu, Y. 2021. Impact of invasive ant species on native fauna across similar habitats under global environmental changes. Environmental Science and Pollution Research 28(39), 54362–54382 (doi:10.1007/s11356-021-15961-5).
- Singh, S., Colombo, W.D., Shreevihar, S., Faisal, M., Pandey, Y,V.P., Tribull, C.M. 2021. Rediscovery of Kurian’s types of Hymenoptera at Forest Research Institute, Dehradun, India with notes on Indian bethylid fauna. Zootaxa 50191: 1–90 (doi:10.11646/zootaxa.5019.1.1).
- Smith, D. R. 1979. Superfamily Formicoidea. Pp. 1323-1467 in: Krombein, K. V., Hurd, P. D., Smith, D. R., Burks, B. D. (eds.) Catalog of Hymenoptera in America north of Mexico. Volume 2. Apocrita (Aculeata). Washington, D.C.: Smithsonian Institution Pr (page 1421, see also)
- Sorvari, J. 2002. Tapinoma melanocephalum (Fabricius, 1793) (Hymenoptera: Formicidae), an imported ant species new to Finland, with observations and a taxonomic note. Entomol. Gaz. 53: 269-270 (page 269, see also)
- Terayama, M., Sunamura, E., Fujimaki, R., Ono, T., Eguchi, K. 2021. A Surprisingly Non-attractiveness of Commercial Poison Baits to Newly Established Population of White-Footed Ant, Technomyrmex brunneus (Hymenoptera: Formicidae), in a Remote Island of Japan. Sociobiology 68, 5898 (doi:10.13102/sociobiology.v68i1.5898).
- Tseng, S.-P. 2020. Evolutionary history of a global invasive ant, Paratrechina longicornis (Dissertation_全文 ). Ph.D. thesis, Kyoto University.
- Tseng, S.-P., Hsu, P.-W., Lee, C.-C., Wetterer, J.K., Hugel, S., Wu, L.-H., Lee, C.-Y., Yoshimura, T., Yang, C.-C.S. 2020. Evidence for common horizontal transmission of Wolbachia among ants and ant crickets: Kleptoparasitism added to the list. Microorganisms 8, 805. (doi:10.3390/MICROORGANISMS8060805).
- Ulysséa, M.A., Brandão, C.R.F. 2013. Ant species (Hymenoptera, Formicidae) from the seasonally dry tropical forest of northeastern Brazil: a compilation from field surveys in Bahia and literature records. Revista Brasileira de Entomologia 57, 217–224 (doi:10.1590/s0085-56262013005000002).
- Varela-Hernández, F., Medel-Zosayas, B., Martínez-Luque, E.O., Jones, R.W., De la Mora, A. 2020. Biodiversity in central Mexico: Assessment of ants in a convergent region. Southwestern Entomologist 454: 673-686.
- Wang, W.Y., Soh, E.J.Y., Yong, G.W.J., Wong, M.K.L., Benoit Guénard, Economo, E.P., Yamane, S. 2022. Remarkable diversity in a little red dot: a comprehensive checklist of known ant species in Singapore (Hymenoptera: Formicidae) with notes on ecology and taxonomy. Asian Myrmecology 15: e015006 (doi:10.20362/am.015006).
- Wetterer, J. K. 2009. Worldwide spread of the ghost ant, Tapinoma melanocephalum (Hymenoptera: Formicidae). Myrmecol. News. 12:23-33.
- Wetterer, J.K. 2021. Ants (Hymenoptera, Formicidae) of St. Vincent, West Indies. Sociobiology 68, e6725 (doi:10.13102/sociobiology.v68i2.6725).
- Wetterer, J.K., Espadaler, X., Ashmole, N.P., Mendel, H., Cutler, C., Endeman, J. 2007. Ants (Hymenoptera: Formicidae) of the South Atlantic islands of Ascension Island, St Helena, and Tristan da Cunha. Myrmecological News 10: 29-37.
- Wheeler, G. C.; Wheeler, J. 1951. The ant larvae of the subfamily Dolichoderinae. Proc. Entomol. Soc. Wash. 53: 169-210 (page 197, larva described)
- Wilson, E. O.; Taylor, R. W. 1967b. The ants of Polynesia (Hymenoptera: Formicidae). Pac. Insects Monogr. 14: 1-109 (page 80, Senior synonym of australis)
- Wu, J. & Wang, C. 1992. Formicidae (pp. 1301-1320). In Peng, J. et al. Iconography of Forest Insects in Hunan, China. Forest Bureau of Hunan Province: 1473 pp. Hunan Scientific and Technical Publishing House.
- Yamane, S., Tanaka, H.O., Hasimoto, Y., Ohashi, M., Meleng, P., Itioka, T. 2021. A list of ants from Lambir Hills National Park and its vicinity, with their biological information: Part II. Subfamilies Leptanillinae, Proceratiinae, Amblyoponinae, Ponerinae, Dorylinae, Dolichoderinae, Ectatomminae and Formicinae. Contributions from the Biological Laboratory, Kyoto University 31, 87–157.
- Yu, Y. 2016. Risk of alien species introduction to Ogasawara Islands : Case study of ants at Tokyo Port. World Heritage Studies 1, 86-89.
- Zhou, A. M., B. Q. Kuang, and Y. R. Gao. 2015. Does the host plant affect the benefits from mutualisms? The invasive mealybug and ghost ant association. Ecological Entomology. 40:782-786. doi:10.1111/een.12258
- Zhou, A.M., Liang, G.W., Zeng, L., Lu, Y.Y., Xu, Y.J. 2014. Interactions between Ghost Ants and invasive mealybugs: The case of Tapinoma melanocephalum (Hymenoptera: Formicidae) and Phenacoccus solenopsis (Hemiptera: Pseudococcidae). Florida Entomologist 97, 1474–1480 (doi:10.1653/024.097.0423).
- Zima, J., Lebrasseur, O., Borovanska, M., Janda, J. 2016. Identification of microsatellite markers for a worldwide distributed, highly invasive ant species Tapinoma melanocephalum (Hymenoptera: Formicidae). European Journal of Entomology 113: 409–414 (doi: 10.14411/eje.2016.053).
References based on Global Ant Biodiversity Informatics
- Abe T., S. Yamane, and K. Onoyama. Ants collected on the Krakatau Islands 100 years after the great eruptions. Biogeography 14: 65-75.
- Alcantara M. J., S. Modi, T. C. Ling, J. Monkai, H. Xu, S. Huang, and A. Nakamura. 2019. Differences in geographic distribution of ant species (Hymenoptera: Formicidae) between forests and rubber plantations: a case study in Xishuangbanna, China, and a global meta-analysis. Myrmecological News 29: 135-145.
- Amarasinghe H. E. 2010. Species composition and nesting habits of ants in a hill-country home garden in Sri Lanka. Asian Myrmecology 3: 9-20.
- André E. 1892. Voyage de M. Chaper à Bornéo. Catalogue des fourmis et description des espèces nouvelles. Mém. Soc. Zool. Fr. 5: 46-55.
- Anh L. N., K. Ogata, and S. Hosoichi. 2010. Ants of agricultural fields in Vietnam (Hymenoptera: Formicidae). Bull. Inst. Trop. Agr., Kyushu Univ. 33: 1-11.
- Baroni Urbani C. 1977. Katalog der Typen von Formicidae (Hymenoptera) der Sammlung des Naturhistorischen Museums Basel (2. Teil). Mitt. Entomol. Ges. Basel (n.s.) 27: 61-102.
- Bharti H., Y. P. Sharma, M. Bharti, and M. Pfeiffer. 2013. Ant species richness, endemicity and functional groups, along an elevational gradient in the Himalayas. Asian Myrmecology 5: 79-101.
- Bharti H., Y. P. Sharma, and A. Kaur. 2009. Seasonal patterns of ants (Hymenoptera: Formicidae) in Punjab Shivalik. Halteres 1(1): 36-47.
- Borowiec L., and S. Salata. 2019. Next step in the invasion: Trichomyrmex mayri (Forel, 1902) new to the Philippines (Hymenoptera: Formicidae). Annals of the Upper Silesian Museum in Bytom Entomology 28(3): 1-3.
- Chapman, J. W., and Capco, S. R. 1951. Check list of the ants (Hymenoptera: Formicidae) of Asia. Monogr. Inst. Sci. Technol. Manila 1: 1-327
- Cheesman L. E., and W. C. Crawley. 1928. A contribution towards the insect fauna of French Oceania. - Part III. Formicidae. Ann. Mag. Nat. Hist. 10(2): 514-525.
- Chen P., Y. Su, S. S. Rao, Y. F. Long, and C. H. Du. 2012. Study on the ant diversity in different Illicium verum stands. Journal of West China Forestry Science 41(1): 60-68.
- Chen Y. Q., Q. Li, Y. L. Chen, Z. X. Lu, X. Y. Zhou. 2011. Ant diversity and bio-indicators in land management of lac insect agroecosystem in Southwestern China. Biodivers. Conserv. 20: 3017-3038.
- Chen Y., C. W. Luo, H. W. Li, Y. J. Liu, H. F. Zheng, and F. C. Yang. 2013. Investigation of ant species and distribution on Wuliang Mountain. Journal of Henan Agricultural Sciences 42(5): 118-122.
- Chen Y., C.-W. Luo, H. W Li, Z. H. Xu, Y. J. Liu, and S. J. Zhao. 2011. The investigation of soil ant resources on the West slope of Mt Ailao. Hubei Agricultural Sciences 50(7): 1356-1359.
- Cheng D., Z. Chen, and S. Zhou. 2015. An analysis on the ant fauna of Jinzhongshan Nature Reserve in Gunagxi, China. Journal of Guangxi Normal University: Natural Science Edition 33(3): 129.137.
- Crawley W.C. 1924. Ants from Sumatra, with biological notes by Edward Jacobson. Annals and Magazine of Natural History (9)13: 380-409
- D'Cunha P., and V. M. Grover Nair. 2014. Ant fauna on the mangroves of Dakshina Kannada and Udupi districts, Karnataka, India. Journal of Ent. Res. 38(1): 59-66.
- Dad J. M., S. A. Akbar, H. Bharti, and A. A. Wachkoo. 2019. Community structure and ant species diversity across select sites ofWestern Ghats, India. Acta Ecologica Sinica 39: 219–228.
- Dhote J. 2012. Ant (Hymenoptera: Formicidae) records from Shri Shivaji science college campus, Amravati, India. Applied Research and Development Institute Journal 3(12): 114-117.
- Dias R. K. S. 2002. Current knowledge on ants of Sri Lanka. ANeT Newsletter 4: 17- 21.
- Dias R. K. S. 2006. Current taxonomic status of ants (Hymenoptera: Formicidae) in Sri Lanka. The Fauna of Sri Lanka: 43-52. Bambaradeniya, C.N.B. (Editor), 2006. Fauna of Sri Lanka: Status of Taxonomy, Research and Conservation. The World Conservation Union, Colombo, Sri Lanka & Government of Sri Lanka. viii + 308pp.
- Dias R. K. S., K. R. K. A. Kosgamage, and H. A. W. S. Peiris. 2012. The Taxonomy and Conservation Status of Ants (Order: Hymenoptera, Family: Formicidae) in Sri Lanka. In: The National Red List 2012 of Sri Lanka; Conservation Status of the Fauna and Flora. Weerakoon, D.K. & S. Wijesundara Eds., Ministry of Environment, Colombo, Sri Lanka. p11-19.
- Dias R. K. S., and K. R. K. A. Kosgamage. Systematics and community composition of foraging workers ants (Family: Formicidae) collected from three habitats in a dry zone region of Sri Lanka. Proceedings of the Annual Research Symposium 2008. Faculty of Graduate Studies, University of Kelaniya.
- Dias R. K. S., and K. R. K. Anuradha Kosgamage. 2012. Occurrence and species diversity of ground-dwelling worker ants (Family: Formicidae) in selected lands in the dry zone of Sri Lanka. J. Sci. Univ. Kelaniya 7: 55-72.
- Dias R. K. S., and R. P. K. C. Rajapaksa. 2016. Geographic records of subfamilies, genera and species of ants (Hymenoptera: Formicidae) in the four climatic zones of Sri Lanka: a review. J. Sci. Univ. Kelaniya 11(2): 23-45.
- Dias R. K. S., and W. S. Udayakantha. 2016. Discovery of the Sri Lankan Relict Ant, Aneuretus simoni Emery (Formicidae, Aneuretinae) and the nest density of the species in a selected region of Meethirigala Forest Reserve, Sri Lanka. Asian Myrmecology 8: 1-8. DOI: 10.20362/am.008005
- Dias R. K. S.; Perera K. A. M. 2011. Worker ant community observed by repeated sampling and information on endemic Aneuretus simoni Emery in the Gilimale Forest Reserve in Sri Lanka. Asian Myrmecology 4: 69-78.
- Dias, R.K.S. 2006. Overview of ant research in Sri Lanka: 2000-2004. ANeT Newsletter 8:7-10
- Emery C. 1893. Voyage de M. E. Simon aux îles Philippines (mars et avril 1890). Formicides. Annales de la Société Entomologique de France 62: 259-270.
- Emery C. 1893. Voyage de M. E. Simon à l'île de Ceylan (janvier-février 1892). Formicides. Annales de la Société Entomologique de France 62: 239-258.
- Emery C. 1901. Ameisen gesammelt in Ceylon von Dr. W. Horn 1899. Deutsche Entomologische Zeitschrift 1901: 113-122.
- Emery C. Formiche raccolte da Elio Modigliani in Sumatra, Engano e Mentawei. Annali del Museo Civico di Storia Naturale 40: 661-722.
- Emery, C. "Formiche raccolte da Elio Modigliani in Sumatra, Engano e Mentawei." Annali del Museo Civico di Storia Naturale Giacomo Doria (Genova) (2) 20, no. 40 (1900): 661-722.
- Fayle T.M., Bakker, L., Cheah, C., Ching, T.M., Davey, A., Dem, F., Earl, A., Huaimei, Y., Hyland, S., Johansson, B., Ligtermoet, E., Lim, R., Lin, L.K., Luangyotha, P., Martins, B.H., Palmeirim, A.F., Paninhuan, S., Rojas, S.K., Sam, L., Sam, P.T.T., Susanto, D., Wahyudi, A., Walsh, J., Weigl, S., Craze, P.G., Jehle, R., Metcalfe, D. & Trevelyan, R. 2011. A positive relationship between ant biodiversity (Hymenoptera: Formicidae) and rate of scavenger-mediated nutrient redistribution along a disturbance gradient in a south-east Asian rain forest. Myrmecological News 14: 5-12.
- Fellowes J. R. 1999. Exotic ants in Asia: is the mainland at risk? The case of Hong Kong. Aliens 9: 5-6.
- Forel A. 1895. Les Formicides de l'Empire des Indes et de Ceylan. Part V. J. Bombay Nat. Hist. Soc. 9: 453-472.
- Forel A. 1911. Ameisen aus Ceylon, gesammelt von Prof. K. Escherich (einige von Prof. E. Bugnion). Pp. 215-228 in: Escherich, K. Termitenleben auf Ceylon. Jena: Gustav Fischer, xxxii + 262 pp.
- Forel A. 1913. H. Sauter's Formosa-Ausbeute: Formicidae II. Arch. Naturgesch. (A)79(6): 183-202
- Forel A. 1913k. Wissenschaftliche Ergebnisse einer Forschungsreise nach Ostindien ausgeführt im Auftrage der Kgl. Preuss. Akademie der Wissenschaften zu Berlin von H. v. Buttel-Reepen. II. Ameisen aus Sumatra, Java, Malacca und Ceylon. Gesammelt von Herrn Prof. Dr. v. Buttel-Reepen in den Jahren 1911-1912. Zoologische Jahrbücher. Abteilung für Systematik, Geographie und Biologie der Tiere 36:1-148.
- General D. M., and G. D. Alpert. 2012. A synoptic review of the ant genera (Hymenoptera, Formicidae) of the Philippines. Zookeys 200: 1-111.
- Ghosh S. N., S. Sheela, B. G. Kundu, S. Roychowdhury, and R. N. Tiwari. 2006. Insecta: Hymenoptera: Formicidae. Pp. 369-398 in: Alfred, J. R. B. (ed.) 2006. Fauna of Arunachal Pradesh. (Part -2). [State Fauna Series 13.]. New Delhi: Zoological Survey of India, iv + 518 pp.
- Ghosh S. N., S. Sheela, and B. G. Kundu. 2005. Ants (Hymenoptera: Formicidae) of Rabindra Sarovar, Kolkata. Records of the Zoological Survey of India. Occasional Paper 234: 1-40.
- Guerrero R. J. 2018. Taxonomic identity of the ghost ant, Tapinoma melanocephalum (Fabricius, 1793) (Formicidae: Dolichoderinae). Zootaxa 4410(3): 497-510.
- Guénard B., and R. R. Dunn. 2012. A checklist of the ants of China. Zootaxa 3558: 1-77.
- Herwina H., N. Nasir, Jumjunidang, and Yaherwandi. 2013. The composition of ant species on banana plants with Banana Bunchy-top Virus (BBTV) symptoms in West Sumatra, Indonesia. Asian Myrmecology 5: 151-161.
- Herwina H., and K. Nakamura. 2007. Ant species diversity study using pitfall traps in a small yard in Bogor Botanic garden, West Java, Indonesia. Treubia 35: 99-116.
- Hosoichi S., S. H. Park, S. Tagane, M. M. Rahman, and K. Ogata. 2018. Domatia of the ant-plant Hydnophytum formicarum (Rubiaceae) captured as nests by two widespread ant species, Tapinoma melanocephalum and Tetramorium nipponense (Hymenoptera: Formicidae). Entomological News 127(5): 407-412.
- Hu C.-H. 2006. Indigenized conservation and biodiversity maintenance on Orchid Island. PhD Thesis, graduate school of the University of Minnesota. 150 pages.
- Imai H. T., M. Kubota, W. L. Brown, Jr., M. Ihara, M. Tohari, and R. I. Pranata. 1985. Chromosome observations on tropical ants from Indonesia. Annu. Rep. Natl. Inst. Genet. Jpn. 35: 46-48.
- Ito, F.; Yamane, S.; Eguchi, K.; Noerdjito, W. A.; Kahono, S.; Tsuji, K.; Ohkawara, K.; Yamauchi, K.; Nishida, T.; Nakamura, K. 2001. Ant species diversity in the Bogor Botanic Garden, West Java, Indonesia, with descriptions of two new species of the genus Leptanilla (Hymenoptera, Formicidae). Tropics 10:379-404.
- Jaitrong W., B. Guenard, E. P. Economo, N. Buddhakala, and S. Yamane. 2016. A checklist of known ant species of Laos (Hymenoptera: Formicidae). Asian Myrmecology 8: 1-32. DOI: 10.20362/am.008019
- Jaitrong W., and T. Ting-Nga. 2005. Ant fauna of Peninsular Botanical Garden (Khao Chong), Trang Province, Southern Thailand (Hymenoptera: Formicidae). The Thailand Natural History Museum Journal 1(2): 137-147.
- Jaitrong W.; Nabhitabhata, J. 2005. A list of known ant species of Thailand. The Thailand Natural History Museum Journal 1(1): 9-54.
- Karmaly K. A.; S. Sumesh, T. P. Rabeesh, and L. Kishore. 2010. A checklist of ants of Thirunelli in Wayanad, Kerala. J. of the Bombay Natural History Society 107(1): 64-67.
- Kishimoto-Yamata K., F. Hyodo, M. Matsuoka, Y. Hashimoto, M. Kon, T. Ochi, S. Yamane, R. Ishii, and T. Itioka. 2012. Effects of remnant primary forests on ant and dung beetle species diversity in a secondary forest in Sarawak, Malaysia. Journal of Insect Conservation DOI 10.1007/s10841-012-9544-6
- Kosgamage K. R. K. A., and R. K. S. Dias 2008. Systematics and community composition of Foraging worker ants ((Order: Hymenoptera, Family: Formicidae) collected from three habitats on a dry zone region of Sri Lanka.Proceedings of Postgraduate Symposium of Kelaniya University. 115pp.
- Lee T. L., and Y. S. Wei. 2005. Study for the temporal and spatial variation of the ant assemblages as the biological indicator in national parks. Journal of Animal and Veterinary Advances 4(4): 491-496.
- Leong C. M., S. F. Shiao, and B. Guenard. 2017. Ants in the city, a preliminary checklist of Formicidae (Hymenoptera) in Macau, one of the most heavily urbanized regions of the world. Asian Myrmecology 9: e009014.
- Li Q., B. D. Hoffmann, Z. X. Lu, and Y. Q. Chen. 2017. Ants show that the conservation potential of afforestation efforts in Chinese valley-type savanna is dependent upon the afforestation method. Journal of Insect Conservation DOI 10.1007/s10841-017-0005-0
- Li Q., Y. Chen, S. Wang, Y. Zheng, Y. Zhu, and S. Wang. 2009. Diversity of ants in subtropical evergreen broadleaved forest in Pu'er City, Yunnan. Biodiversity Science 17(3): 233-239.
- Li Q., Z. Lu, Z. Wei, M. Yanyan, and F. Ping. 2015. Communities of ground-dwelling ants in different plantation forest in arid-hot valleys of Jinsha river, Yunnan Province, China. Scientia Silvae Sinicae 51(8): 134-142.
- Li Z.h. 2006. List of Chinese Insects. Volume 4. Sun Yat-sen University Press
- Liu X. 2012. Taxonomy, diversity and spatial distribution characters of the ant family Formicidae (Insecta: Hymenoptera) in southeastern Tibet. PhD Thesis 139 pages
- Lopwichan S., and S. Khachonpisitsak. 2015. Ant diversity in Nong Tha Yu Arboretum, Si Racha District, Chon Buri Province. Proceedings The 7 th National Science Research Conference. 30-31 March 2015. Naresuan University.
- Lu Z. X., Y. Q. Chen, Q. Li, S. M. Wang, C. J. Liu, and W. Zhang. 2012. Effect of lac insect honeydew on the diversity of ground-dwelling ants in lac plantation. Chin. J. Appl. Ecol. 23(4): 1117-1122.
- Lu Z., K. Li, N. Zhang, and Y. Chen. 2017. Diversity and indicator species of leaf-litter ants in Eucalyptus grandis plantations and secondary natural forests. Forest Research 29(4): 576-580
- Lu Z., Y. Chen, Q. Li, S. Wang, C. Liu, and W. Zhang. 2012. Effect of population of Kerria yunnanensis on diversity of ground dwelling ant. Acta Ecologica Sinica 32(19): 6195-6202.
- Lu Z., Y. Chen, W. Zhang, S. Wang, and Q. Li. 2013. Effects of facultative mutualism between ants and lac insects on the diversity of ant communities. Biodiversity Science 21(3): 343-351.
- Lu Z., and Y. Chen. 2016. Effects of habitat on ant functional groups: a case study of Luchun County, Yunnan Province, China. Chinese Journal of Eco-Agriculture 24(5): 801-810.
- Mezger D., and M. Pfeiffer. 2011. Influence of the arrival of Anoplolepis gracilipes (Hymenoptera: Formicidae) on the composition of an ant community in a clearing in Gunung Mulu National Park, Sarawak, Malaysia. Asian Myrmecology 4: 89-98.
- Mohanraj P., M. Ali, and K. Veerakumari. 2010. Formicidae of the Andaman and Nicobar Islands (Indian Ocean: Bay of Bengal). Journal of Insect Science 10: Article 172
- Mohanraj, P., M. Ali and K. Veenakumari. 2010. Formicidae of the Andaman and Nicobar Islands (Indian Ocean: Bay Of Bengal). Journal of Insect Science 10:172.
- Mukherji D., and S. Ribeiro. 1925. On a collection of ants (Formicidae) from the Andaman Islands. Records of the Indian Museum 27: 205-209.
- Musthak Ali T. M. 1982. Ant fauna (Hymenoptera: Formicidae) of Bangalore with observations on their nesting and foraging habits. Thesis Abstracts. Haryana Agricultural University 8: 370-371.
- Ngoc Anh L., K. Ogata, and S. Hosoishi. 2010. Ants of agricultural fields in Vietnam (Hymenoptera: Formicidae). Bull. Inst. Trop. Agr. Kyushu Univ. 33: 1-11.
- Nie L., M. Ni, D. Ning, H. Ran, B. Hassan, and Y. Xu. 2019. Comparing mechanisms of competition among introduced and resident ants in China: from behavior to trophic position (Hymenoptera: Formicidae). Myrmecological News 29: 125-133.
- Pajni H. R., and R. K. Suri. 1978. First report on the Formicid fauna (Hymenoptera) of Chandigarh. Res. Bull. (Science) Punjab University 29: 5-12.
- Parui A. K., S. Chatterjee, and P. Basu. 2015. Habitat characteristics shaping ant species assemblages in a mixed deciduous forest in Eastern India. Journal of Tropical Ecology: 1-14. doi:10.1017/S0266467415000036
- Pfeiffer M., D. Mezger, and J. Dyckmans. 2013. Trophic ecology of tropical leaf litter ants (Hymenoptera: Formicidae) - a stable isotope study in four types of Bornean rain forest. Myrmecological News 19: 31-41.
- Pfeiffer M., and D. Mezger. 2012. Biodiversity Assessment in Incomplete Inventories: Leaf Litter Ant Communities in Several Types of Bornean Rain Forest. PLoS ONE 7(7): e40729. doi:10.1371/journal.pone.0040734
- Pfeiffer M., and D. Mezger. 2012. Biodiversity Assessment in Incomplete Inventories: Leaf Litter Ant Communities in Several Types of Bornean Rain Forest. PLoS ONE 7(7): e40729. doi:10.1371/journal.pone.0040846
- Pfeiffer M.; Mezger, D.; Hosoishi, S.; Bakhtiar, E. Y.; Kohout, R. J. 2011. The Formicidae of Borneo (Insecta: Hymenoptera): a preliminary species list. Asian Myrmecology 4:9-58
- Pfeiffer, M., H. Cheng Tuck, and T. Chong Lay. 2008. Exploring arboreal ant community composition and co-ccurrence patterns in plantations of oil palm Elaeis guineensis in Borneo and Peninsular Malaysia. Ecography 31(1): 21-32.
- Philpott S.M., P. Bichier, R.A. Rice, and R. Greenberg. 2008. Biodiversity conservation, yield, and alternative products in coffee agroecosystems in Sumatra, Indonesia. Biodivers. Conserv. 17: 1805-1820. Data obtained from Stacy Philpott
- Raci N., C. Sravanthy, C. Sammaiah, and M. Thirupahaiah. 2015. Biodiversity of ants (Insecta-Hymenoptera) in agroecosystem and grass land in Jammikunta, Karimnagar District, Telangana, India. Journal ofEnvironment 4(1): 11-16.
- Rajagopal T., S. P. Sevarkodiyone, and A. Manimozhi. 2005. Ant diversity in some selected localities of Sattur Taluk, Virudhunagar district, Tamil Nadu. Zoos' Print Journal 20(6): 1887-1888.
- Rajan P. D., M. Zacharias, and T. M. Mustak Ali. 2006. Insecta: Hymenoptera: Formicidae. Fauna of Biligiri Rangaswamy Temple Wildlife Sanctuary (Karnataka). Conservation Area Series, Zool. Surv. India.i-iv,27: 153-188.
- Ramesh T., K. J. Hussain, K. K. Satpathy, M. Selvanayagam, and M. V. R. Prasad. 2010. Diversity, distribution and species composition of ants fauna at Department of Atomic Energy (DAE) Campus Kalpakkam, South India. World Journal of Zoology 5 (1): 56-65.
- Rizali A., A. Rahim, B. Sahari, L.B. Prasetyo, and D. Buchori. 2011. Impact of invasive ant species in shaping ant community structure on small islands in Indonesia. Jurnal Biologi Indonesia 7(2): 221-230.
- Rizali A., M. M. Bos, D. Buchori, Sk. Yamane, and C. H. Schulze. 2008. Ants in tropical urban habitats: the myrmecofauna in a densely populated area of Bogor, West Java, Indonesia. Hayati Journal of Biosciences 15(2): 77-84.
- Rizali A., M.M. Bos, D. Buchori, Sk. Yamane, C. Hans, and J. Schulze. 2008. Ants in tropical urban habitats: the myrmecofauna in a densely populated area of Bogor, West Java, Indonesia. Hayati Journal of Biosciences 77-84.
- Rizali A., Y. Clough, D. Buchori, M. L . A. Hosang, M. M. Bos, and T. Tscharntke. 2012. Long-term change of ant community structure in cacao agroforestry landscapes in Indonesia. Insect Conservation and Diversity doi: 10.1111/j.1752-4598.2012.00219.x
- Sakchoowong W., W. Jaitrong, and K. Ogata. 2009. Comparison of ground-ant diversity between natural forests and disturbed forests along a natural gas pipeline transect in Thong Pha Phum National Park, Kanchanaburi province. Kasetsart J. (Nat. Sci) 43: 64-73.
- Savitha S., N. Barve, and P. Davidar. 2008. Response of ants to disturbance gradients in and around Bangalore, India. Tropical Ecology 49(2): 235-243.
- Shattuck S. O. 1994. Taxonomic catalog of the ant subfamilies Aneuretinae and Dolichoderinae (Hymenoptera: Formicidae). University of California Publications in Entomology 112: i-xix, 1-241.
- Shukla R. K., H. Singh, N. Rastogi, and V. M. Agarwal. 2013. Impact of abundant Pheidole ant species on soil nutrients in relation to the food biology of the species. Applied Soil Ecology 71: 15 23.
- Sitthicharoenchai D., and N. Chantarasawat. 2006. Ant species diversity in the establishing area for Advanced Technology Institute at Lai-Nan Sub-district, Wiang Sa district, Nan Province, Thailand. The Natural History Journal of Chulalongkorn University 6(2): 67-74
- Skarbek C. J., M. Noack, H. Bruelheide, W. Hardtle, G. von Oheimb, T. Scholten, S. Seitz, M. Staab. 2019. A tale of scale: plot but not neighbourhood tree diversity increases leaf litter ant diversity. Journal of Animal Ecology DOI: 10.1111/1365-2656.13115
- Song Y., Z. H. Xu, C. L. Li, Y. Q. Hao, and H. B. Li. 2014. Ant communities of Nangun River Nature Reserve in Yunnan. Journal of West China Forestry Science 45(5): 93-100.
- Song Y., Z. Xu, C. Li, N. Zhang, L. Zhang, H. Jiang, and F. Mo. 2013. An Analysis on the Ant Fauna of the Nangun river Nature Reserve in Yunnan, China. Forest Research 26(6): 773-780.
- Sonune B. V., and R. J. Chavan. 2016. Distribution and diversity of ants (Hymenoptera: Formicidae) around Gautala Autramghat Sanctuary, Aurangabad Maharashtra, India. Journal of Entomology and Zoology Studies 4(2): 361-364.
- Staab M., J. Methorst, J. Peters, N. Bluthgen, and A. M. Klein. 2017. Tree diversity and nectar composition affect arthropod visitors on extrafloral nectaries in a diversity experiment. Journal of Plant Ecology 10(1): 210-212.
- Staab M., N. Bluthgen, and A. M. Klein. 2014. Tree diversity alters the structure of a tri-trophic network in a biodiversity experiment. Oikos doi: 10.1111/oik.01723
- Tak N. 1995. Studies on ants (Formicidae) of Rajasthan - 1 Jodhpur. Hexapoda 7(1): 17-28.
- Tak N. 2000. Studies on ants (Formicidae) of Rajasthan--II Dungarpur. Entomon 25: 47-54.
- Tak N. 2008. Ants of Rajasthan. Conserving Biodiversity of Rajasthan Zool. Surv. India. 149-155.
- Tak N. 2009. Ants Formicidae of Rajasthan. Records of the Zoological Survey of India, Occasional Paper No. 288, iv, 46 p
- Tak N. 2010. Insecta: Hymenoptera: Formicidae. Zool. Surv. India, Fauna of Ranthambore National Park, Conservation Area Series 43: 133-144.
- Tak N., N. S. Rathore, and S. Kumar. 2007. Insecta: Hymenoptera. Fauna of Pichhola lake (Rajasthan). Wetland Ecosystem series. Zool. Surv. India. 8 : 127-130.
- Tak N., and N. S. Rathore. 1996. Ant (Formicidae) fauna of the Thar Desert. Pp. 271-276 in: Ghosh, A. K.; Baqri, Q. H.; Prakash, I. (eds.) 1996. Faunal diversity in the Thar Desert: gaps in research. Jodhpur: Scientific Publishers, xi + 410 pp.
- Tak N., and N. S. Rathore. 2004. Insecta: Hymenoptera: Formicidae. State Fauna Series 8: Fauna of Gujarat. Zool. Surv. India. Pp. 161-183.
- Tak N., and S. I. Kazmi. 2013. On some ants (Insecta: Hymenoptera: Formicidae) from Nagaland, India. Rec. zool. Surv. India: 113(1): 169-182.
- Tak N., and S. L. Kazmi. 2011. On a collection of Insecta: Hymenoptera: Formicidae from Uttarakhand. Rec. zool. Surv. India : 111(2) : 39-49.
- Tak, N. 2009. Ants (Hymenoptera: Formicidae) of the Thar Desert of Rajasthan and Gujarat. in C. Sivaperuman et al. (eds.), Faunal Ecology and Conservation of the Great Indian Desert
- Terayama M. 2009. A synopsis of the family Formicidae of Taiwan (Insecta: Hymenoptera). Research Bulletin of Kanto Gakuen University. Liberal Arts 17:81-266.
- Terayama, M. 2009. A synopsis of the family Formicidae of Taiwan (Insecta; Hymenoptera). The Research Bulletin of Kanto Gakuen University 17: 81-266.
- Trainor C.R. and A.N. Andersen. 2010. The ant fauna of Timor and neighbouring islands: potential bridges between the disjunct faunas of South East Asia and Australia. Australian Journal of Zoology 58: 133-144.
- Trainor C.R., and A.N. Andersen. 2010. The ant fauna of Timor and neighbouring islands: potential bridges between the disjunct faunas of South East Asia and Australia. Australian Journal of Zoology 58: 133-144.
- Varghese T. 2004. Taxonomic studies on ant genera of the Indian Institute of Science campus with notes on their nesting habits. Pp. 485-502 in : Rajmohana, K.; Sudheer, K.; Girish Kumar, P.; Santhosh, S. (eds.) 2004. Perspectives on biosystematics and biodiversity. Prof. T.C. Narendran commemoration volume. Kerala: Systematic Entomology Research Scholars Association, xxii + 666 pp.
- Wang C. and Wu J. 1992. Ants of the Jianfengling forest region in Hainan Province (Hymenoptera: Formicidae). Scientia Silvae Sinicae 28: 561-564.
- Wang W. R., S. Q. Zhang, and Z. H. Xu. 2012. A faunistic and taxonomic study of ants (Hymenoptera: Formicidae) in Shenzhen Municipality. Journal of Southwest Forestry University 32(1): 64-73.
- Wang W., S. Zhang, and Z Xu. 2012. Distribution patters of ant species in Shenzhen City. Journal of Southwest Forestry University 32(3): 70-74.
- Wang W., S. Zhang, and Z. Xu. 2012. Distribution Patterns of Ant Species in Shenzhen City. Journal of Southwest Forestry University 32(3): 69-74.
- Watanasit S., S. Sonthichai, and N. Noon-anant. 2003. Preliminary survey of ants at Tarutao National Park, Southern Thailand. Songklanakarin J. Sci. Technol. 25(1) : 115-122
- Way M. J., and B. Bolton. 1997. Competition between ants for coconut palm nesting sites. Journal of Natural History 31: 439-455.
- Wheeler W. M. 1909. Ants of Formosa and the Philippines. Bulletin of the American Museum of Natural History 26: 333-345.
- Wheeler W. M. 1913. Zoological results of the Abor Expedition, 1911-1912, XVII. Hymenoptera, II: Ants (Formicidae). Records of the Indian Museum 8: 233-237.
- Wheeler W. M. 1919. The ants of Borneo. Bulletin of the Museum of Comparative Zoology 63:43-147.
- Wheeler W. M. 1923. Chinese ants collected by Professor S. F. Light and Professor A. P. Jacot. American Museum Novitates 69: 1-6.
- Wheeler W. M. 1927. Ants collected by Professor F. Silvestri in Indochina. Bollettino del Laboratorio di Zoologia Generale e Agraria della Reale Scuola Superiore d'Agricoltura. Portici 20: 83-106.
- Wheeler W. M. 1928. Ants collected by Professor F. Silvestri in China. Bollettino del Laboratorio di Zoologia Generale e Agraria della Reale Scuola Superiore d'Agricoltura. Portici 22: 3-38.
- Wheeler W. M. 1929. Ants collected by Professor F. Silvestri in Formosa, the Malay Peninsula and the Philippines. Bollettino del Laboratorio di Zoologia Generale e Agraria della Reale Scuola Superiore d'Agricoltura. Portici 24: 27-64.
- Wheeler W. M. 1929. Some ants from China and Manchuria. American Museum Novitates 361: 1-11.
- Wheeler W. M. 1930. A list of the known Chinese ants. Peking Natural History Bulletin 5: 53-81.
- Wheeler W. M. 1930. Formosan ants collected by Dr. R. Takahashi. Proceedings of the New England Zoological Club 11: 93-106.
- Wheeler W. M. 1937. Additions to the ant-fauna of Krakatau and Verlaten Island. Treubia 16: 21-24.
- Wu B., Y. Lu, G. Liang, and L. Zeng. 2010. Influence of the red inported fire ant, Solenopsis invicta Buren (Hymenoptera: Formicidae) on the diversity of ant communities in a newly infested longan orchard and grass areas nearby. Acta Ecologica Sinica 30(8): 2075-2083.
- Wu B., Y. Lu, L. Zeng, and G. Liang. 2008. Influences of Solenopsis invicta Buren invasion on the native ant communities in different habitats in Guangdong. Chin. J. Appl. Ecol. 19(1): 151-156.
- Wu Z. W., X. D. Bi, X. He, Z. X. Lu, L. J. Wei, and Y. Q Chen. 2015. Impact of continuous fire disturbance on ground-dwelling ant communities in arid-hot valleys of Panzhihua, Sichuan. Journal of Yunnan University 37(3): 467-474.
- Xu F. F., and J. Chen. 2009. Comparison of the Differences in Response to the Change of the Extrafloral Nectar-ant-herbivore Interaction System Between a Native and an Introduced Passiflora Species. Acta Botanica Yunnanica 31(6): 543-550.
- Xu Z. H., B. L. Yang, and G. Hu. 1999. Formicidae ant communities in fragments of montane rain forest in Xishuangbanna, China. Zoological Research 20(4): 288-293.
- Xu Z., Zeng G., Liu T.-Y. and He Y.-F. 1999. [A study on communities of Formicidae ants in different subtypes of vegetation in Xishuangbanna District of China.] Zoological Research 20: 118-125
- Yamane S. 2013. A Review of the ant fauna of the Krakatau Islands, Indonesia. Bull. Kitakyushu Mus. Nat. Hist. Hum. Hist. Ser: A, 11: 1-66
- Yamane Sk. 2005. Krakatau in 1982, and the commencement of myrmecological research. The nature and Insects (Konchu to shizen) 40: 27-33.
- Yasumatsu, K. 1940. Matériaux pour servir à la faune myrmécologique des îles de Yaeyama. Mushi 13: 67-70
- Zhang C., Z. Xu, N. Yu, Q. He, and X. Liu. 2012. Distribution Patterns of Ant Species on East Slope of Mount Demola and Zayu Valley in Southeastern Tibet. Journal of Northeast Forestry University 40(3): 87-92
- Zhang N. N., Y. Q. Chen, Z. X. Lu, W. Zhang, and K. L. Li. 2013. Species diversity, community structure difference and indicator species of leaf-litter ants in rubber plantations and secondary natural forests in Yunnan, southwestern China. Acta Entomologica Sinica 56(11): 1314-1323.
- Zhang R. J., L. W. Liang, and S. Y. Zhou. 2014. An analysis on the ant fauna of Nonggang Nature Reserve in Guangxi, China. Journal of Guangxi Normal university: Natural Science Edition 32(3): 86-93.
- Zhang W., G. Liu, P. Zhong, and S. Zhang. 2014. Investigation of Formicidae in Luofushan Mountain. Journal of Huizhou University 34(3): 46-50.
- Zhang W., and S. Zhou. 2016. An investigation on Formicidae species of Nanling National Park. Journal of Huizhou University 36(3): 27-30.
- Zhang Z., M. Cao, X. Yang, X. Deng, and Y. She. 2000. A study on species diversity of ant in fragments of seasonal rain forest of Xishuangbanna, China. Zoological Research 21(1): 70-75.
- Zhou S.-Y. 2001. Ants of Guangxi. Guangxi Normal University Press, Guilin, China, Guilin, China. 255 pp.
- Zryanin V. A. 2011. An eco-faunistic review of ants (Hymenoptera: Formicidae). In: Structure and functions of soil communities of a monsoon tropical forest (Cat Tien National Park, southern Vietnam) / A.V. Tiunov (Editor). – M.: KMK Scientific Press. 2011. 277 р.101-124.
- Zryanin V. A., and M. V. Mokrousov. 2015. Contribution to the ant fauna of Lombok Island. Proceedings of the 10th ANeT International Conference, 23-26 October 2015, University of Kelaniya, Sri Lanka. 34
- Pages using DynamicPageList3 parser function
- Common Name
- Highly invasive
- Polygynous
- Supercolonies
- Photo Gallery
- North subtropical
- Tropical
- Butterfly Associate
- Host of Lampides boeticus
- Aphid Associate
- Host of Aphis eugeniae
- Host of Aphis fabae
- Host of Aphis fabae solanella
- Host of Aphis gossypii
- Host of Aphis pomi
- Host of Greenidea (Greenidea) decaspermi
- Host of Greenidea (Greenidea) ficicola
- Host of Greenidea (Trichosiphum) formosana
- Host of Hysteroneura setariae
- Host of Macrosiphoniella sanborni
- Host of Myzus persicae
- Host of Ovatus crataegarius
- Host of Pentalonia nigronervosa
- Host of Tinocallis kahawaluokalani
- Host of Tuberolachnus salignus
- Karyotype
- Species
- Extant species
- Formicidae
- Dolichoderinae
- Tapinoma
- Tapinoma melanocephalum
- Dolichoderinae species
- Tapinoma species
- Ssr