Brachyponera sennaarensis
| Brachyponera sennaarensis | |
|---|---|

| |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Arthropoda |
| Class: | Insecta |
| Order: | Hymenoptera |
| Family: | Formicidae |
| Subfamily: | Ponerinae |
| Tribe: | Ponerini |
| Genus: | Brachyponera |
| Species group: | lutea |
| Species: | B. sennaarensis |
| Binomial name | |
| Brachyponera sennaarensis (Mayr, 1862)
| |
| Subspecies | |
| |
| Synonyms | |
| |


This is the only species of subfamily Ponerinae that eats seeds in addition to carnivorous habits (Lévieux & Diomande 1978, Dejean & Lachaud 1994). Widely distributed throughout sub-Saharan Africa, where it inhabits savannas and open forests; it is also the most common member of Ponerinae in southern Arabia. While it is regarded as native at least throughout Africa, it also notably prefers man-impacted habitats, such as human settlements, rubbish dumps and waste ground. Thus it is in question whether the species is indigenous to the Socotra Archipelago. It is a general scavenger but will attack other insects and has a painful sting. Allergic reactions to the sting, sometimes severe, are a problem locally in Arabia (DIB 1992, RrzK et al. 1998), where it is called the "Samsun ant". Probably because of awareness of the painful sting, Socotri people refer to this ant by a specific denomination ("diftim"), as different to the word for ant ("nimihil") (Collingwood et al. 2004)
Pashaei Rad et al. (2018) found this species in Iran in a very low rainfall area in a damp habitat in a house.
| At a Glance | • Facultatively polygynous • Tandem running • Invasive |
Identification
Sharaf et al. (2017) - Worker. Large species (TL 5–6). Head broader than mesosoma, with convex sides and emarginated posterior margin; eyes relatively large; metanotal groove deeply impressed; petiole a high and thick node with a straight anterior surface and a convex posterior surface; first and second gastral tergites separated by a distinct constriction characteristic for ponerine ants; gaster ending with a powerful sting. Dark brown to blackish brown, antennae, tibiae and tarsi reddish. All body surfaces covered with fine and dense pubescence.
Keys including this Species
Distribution
Latitudinal Distribution Pattern
Latitudinal Range: 37.8506133° to -27.1°.
| North Temperate |
North Subtropical |
Tropical | South Subtropical |
South Temperate |
- Source: AntMaps
Distribution based on Regional Taxon Lists
Afrotropical Region: Benin, Burkina Faso, Cameroun, Chad, Eritrea, Gambia, Guinea, Ivory Coast, Kenya, Mali, Namibia, Niger, Nigeria, Saudi Arabia, Senegal, Sierra Leone, Socotra Archipelago, Sudan (type locality), United Arab Emirates, Yemen, Zambia, Zimbabwe.
Oriental Region: India.
Palaearctic Region: Bahrain, Iran, Oman, Qatar.
Distribution based on AntMaps
Distribution based on AntWeb specimens
Check data from AntWeb
Countries Occupied
| Number of countries occupied by this species based on AntWiki Regional Taxon Lists. In general, fewer countries occupied indicates a narrower range, while more countries indicates a more widespread species. |

|
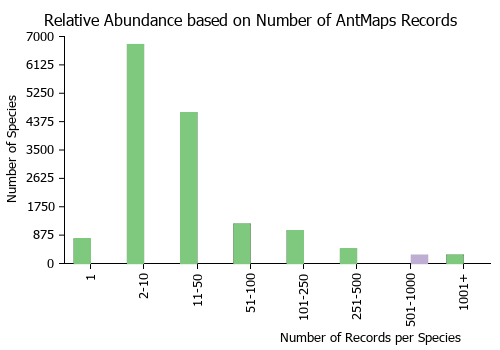
Estimated Abundance
| Relative abundance based on number of AntMaps records per species (this species within the purple bar). Fewer records (to the left) indicates a less abundant/encountered species while more records (to the right) indicates more abundant/encountered species. |
|
Biology
This appears to be the only ponerine ant that can feed on seeds. Diet varies seasonally and geographically: in humid tropical regions of Africa, both seeds and insect prey are collected during the rainy season, whereas diet consists exclusively of seeds during the rainy season that lasts three months (Dejean & Lachaud 1994). In dry tropical regions however, foragers react to the absence of seeds in the rainy season by adopting a 100% animal diet (Lévieux 1979).
Lachaud & Déjean(1994) found that the great flexibility of the feeding strategies exhibited by the ponerine ant Brachyponera senaarensis allows it to exploit either seeds or animal prey items as food resources. Predation is generally limited to small prey and is very similar to scavenging behavior. In laboratory conditions, the predatory behavior of B. senaarensis is not different in structure from that known in other carnivorous ants species. The workers forage individually and return to the nest using a series of cues involving light, a chemical graduated marking system near the nest entrance, and memory. During nest-moving, recruitment by tandem running was observed. However, in colonies where the food supply is regular, workers that discover food do not recruit nestmates, but make repeated trips between the nest and the food source. On the contrary, in starved colonies, the introduction of prey may produce a massive exit of foragers, corresponding to a primitive form of mass recruitment similar to that observed in some other ant species.
Morphology
Billen and Al-Khalifa (2015) - Abstract: The pro- and postpharyngeal glands of Brachyponera sennaarensis both appear as globular formations at a general anatomical level. However, only for the propharyngeal gland do these formations correspond with spherical secretory cells with diameters of 30-40 μm. For the lobed postpharyngeal gland, in contrast, this globular appearance is caused by the bulbous protrusions of the epithelial cells. This lobed appearance and globular cell shape also occur in the postpharyngeal glands of other Ponerinae and thus may represent a phylogenetic character. At the ultrastructural level, the propharyngeal gland cells are characterized by a well-developed granular endoplasmic reticulum, which is in agreement with its presumed production of digestive enzymes. The postpharyngeal gland cells contain a well-developed smooth endoplasmic reticulum, which allows the production of a nonproteinaceous secretion.
Billen and Al-Khalifa (2018) - As in other stinging ants, the venom gland of both queens and workers is formed by two free filaments in which the initial venom synthesis starts, a reservoir sac that encloses the convoluted gland portion, and the venom duct, that opens through the sting (Fig. 1). The paired filaments have a length of approx. 1.5-2 mm and a diameter between 40 and 50 μm, and are situated in between the other abdominal organs with a close proximity to the fat body (Fig. 2A). Both filaments fuse together to become a single unpaired filament shortly before entering the reservoir sac. The unpaired filament is considerably thinner than the paired filaments and has a diameter of 10-15 μm (Fig. 2B). Upon entering the reservoir sac, it expands to form the convoluted gland portion, which is entirely invaginated inside the reservoir sac (Fig. 2C, 4A). The reservoir sac is surrounded by a thick muscular supply (Fig. 2C), while also an occasional nerve can be observed (Fig. 2B). The elongated reservoir sac has a length of approx. 700 μm and a width around 400 μm, and has a cuticle-lined epithelial wall with a thickness of less than 1 μm (Fig. 2D). The venom duct has a length of approx. 750 μm and a diameter around 50 μm and enters the sting base dorsally to the Dufour gland duct, both ducts are associated with bundles of muscle fibres (Fig. 2E,F).
The venom gland filaments consist of class-3 secretory cells (classification of Noirot & Quennedey 1974) that are arranged around a central cuticle-lined filament lumen, into which they open through their accompanying duct cells (Fig. 3A). Each duct cell connects with a secretory cell through the end apparatus, which is formed by a cuticular continuation of the ductule that is surrounded by a sheath of microvilli (Fig. 3B). The cuticular characteristics in the duct cell and the end apparatus differ considerably: in the duct cell, the cuticular lining is thick and continuous, whereas in the end apparatus it is thin and discontinuous (Fig. 3B). The cytoplasm contains a well-developed Golgi apparatus and numerous mitochondria. Endoplasmic reticulum could not be observed, but numerous free ribosomes and polysomes give the cytoplasm a fine granulated appearance (Fig. 3C). In between the secretory cells of the free filaments, tracheoles (not shown) and nerve fibres can be observed (Fig. 3D).
The ovoid convoluted gland is completely internalized inside the reservoir sac, and measures approx. 400x200 μm. Careful tearing open of the reservoir sac of critical point dried material allows observation of the convoluted gland with scanning microscopy (Fig. 4A). Its external surface appears striated, with irregularly scattered small pores with a diameter around 0.5 μm (Fig. 4B). The internal reservoir wall, in contrast, shows a less dense striation without any pores (Fig. 4C). As a continuation of the venom gland filaments, the bulk of the convoluted gland is made up of secretory tissue with class-3 gland cells with a clear end apparatus and their accompanying ducts. The ducts open both along the continuation of the internal lumen inside the convoluted gland lumen and along the periphery of the convoluted gland (Fig. 1, 4D,E).
 Billen and Al-Khalifa (2015) - Figure 1. Scanning electron micrographs of the pharyngeal glands of Brachyponera sennaarensis workers. A. Frontal view of the head with dissected pharyngeal glands (enlarged in B) shown at the same magnification. B. Ventral view of pharynx (ph) with attached pro-(ProPG) and postpharyngeal glands (PPG). C. Detail of secretory cells (SC) and slender duct cells (DC) of propharyngeal gland. Arrows indicate local widening of duct cells in region where duct cell nucleus occurs.
Billen and Al-Khalifa (2015) - Figure 1. Scanning electron micrographs of the pharyngeal glands of Brachyponera sennaarensis workers. A. Frontal view of the head with dissected pharyngeal glands (enlarged in B) shown at the same magnification. B. Ventral view of pharynx (ph) with attached pro-(ProPG) and postpharyngeal glands (PPG). C. Detail of secretory cells (SC) and slender duct cells (DC) of propharyngeal gland. Arrows indicate local widening of duct cells in region where duct cell nucleus occurs. Billen and Al-Khalifa (2018) - Figure 2. A. Scanning micrograph of the sting apparatus with attached hindgut (HG) and Malpighian tubules (MT) of B. sennaarensis worker. The venom gland consists of two free filaments (VF), a reservoir (VGr) with an internalized convoluted gland (visible in 1C) and a venom duct (VD) that opens through the sting (St). B. Detail of framed area in 1A, showing how both filaments fuse into a considerably thinner unpaired filament prior to entering the reservoir sac (white arrow). C. Semithin section through venom reservoir and internalized convoluted gland (CG) in B. sennaarensis alate queen. Note conspicuous supply of muscle fibres (MF). D. Ultrastructural detail (worker) of very thin cuticle-lined reservoir wall. E,F. Longitudinal and transverse semithin sections through the sting base of a worker, showing opening of venom gland duct (VD) and Dufour gland duct (DD). ct: cuticle, F: furcula, FC: fat cells, GS: gonostyli, MF: muscle fibres, NF: nerve fibre, SL: sting lancets.
Billen and Al-Khalifa (2018) - Figure 2. A. Scanning micrograph of the sting apparatus with attached hindgut (HG) and Malpighian tubules (MT) of B. sennaarensis worker. The venom gland consists of two free filaments (VF), a reservoir (VGr) with an internalized convoluted gland (visible in 1C) and a venom duct (VD) that opens through the sting (St). B. Detail of framed area in 1A, showing how both filaments fuse into a considerably thinner unpaired filament prior to entering the reservoir sac (white arrow). C. Semithin section through venom reservoir and internalized convoluted gland (CG) in B. sennaarensis alate queen. Note conspicuous supply of muscle fibres (MF). D. Ultrastructural detail (worker) of very thin cuticle-lined reservoir wall. E,F. Longitudinal and transverse semithin sections through the sting base of a worker, showing opening of venom gland duct (VD) and Dufour gland duct (DD). ct: cuticle, F: furcula, FC: fat cells, GS: gonostyli, MF: muscle fibres, NF: nerve fibre, SL: sting lancets. Billen and Al-Khalifa (2018) - Figure 3. A. Longitudinal semithin section through venom gland filament in worker with central lumen and sections through end apparatus (black arrowheads). B. Electron micrograph detail of end apparatus in worker. Note abrupt transition (arrowheads) of thick and continuous cuticle in duct cell (DC) into thin and discontinuous cuticle in secretory cell (SC). Small white arrows indicate disrupted epicuticular inner lining. C. Ultrastructure of secretory cell cytoplasm (queen) with granular appearance because of abundant free ribosomes, and with well-developed Golgi apparatus (GA) and mitochondria (M). D. Detail of nerve fibre (NF) wedged in between gland cells (queen). mv: microvilli, N: nucleus.
Billen and Al-Khalifa (2018) - Figure 3. A. Longitudinal semithin section through venom gland filament in worker with central lumen and sections through end apparatus (black arrowheads). B. Electron micrograph detail of end apparatus in worker. Note abrupt transition (arrowheads) of thick and continuous cuticle in duct cell (DC) into thin and discontinuous cuticle in secretory cell (SC). Small white arrows indicate disrupted epicuticular inner lining. C. Ultrastructure of secretory cell cytoplasm (queen) with granular appearance because of abundant free ribosomes, and with well-developed Golgi apparatus (GA) and mitochondria (M). D. Detail of nerve fibre (NF) wedged in between gland cells (queen). mv: microvilli, N: nucleus. Billen and Al-Khalifa (2018) - Figure 4. Convoluted gland of B. sennaarensis worker. A. Scanning micrograph showing striated appearance of convoluted gland (CG) after tearing open venom reservoir (RW: reservoir wall). B. Detail of convoluted gland surface (of area indicated by black frame in 4A), arrowheads show openings of gland ducts. C. Detail of inner reservoir wall (of area indicated by white frame in 4A). D,E. Electron micrographs of central (D) and peripheral (E) portion of convoluted gland, showing presence of secretory cells with end apparatus (EA). M: mitochondria, MF: muscle fibres, N: nucleus, VF: venom filaments.
Billen and Al-Khalifa (2018) - Figure 4. Convoluted gland of B. sennaarensis worker. A. Scanning micrograph showing striated appearance of convoluted gland (CG) after tearing open venom reservoir (RW: reservoir wall). B. Detail of convoluted gland surface (of area indicated by black frame in 4A), arrowheads show openings of gland ducts. C. Detail of inner reservoir wall (of area indicated by white frame in 4A). D,E. Electron micrographs of central (D) and peripheral (E) portion of convoluted gland, showing presence of secretory cells with end apparatus (EA). M: mitochondria, MF: muscle fibres, N: nucleus, VF: venom filaments.





Regional Accounts
Oman
Sharaf et al (2018) - This species was found nesting under a stone and foraging on the ground. Several individuals were found in moist soil under a stone next to a date palm tree. In the KSA it has been observed inhabiting sites near human settlements and has an apparent preference of hot habitats (Sharaf & Aldawood, unpublished data)
Saudi Arabia
Al-Khalifa et al (2015) - Collingwood (1985) observed and reported B. sennaarensis in the Kingdom of Saudi Arabia, where it is commonly known as the samsum ant. Later, Collingwood and Agosti (1996) followed and recorded their occurrence in Oman, Yemen and Kuwait, whilst Collingwood et al. (1997) reported it in the United Arab Emirates. B. sennaarensis is considered to constitute a public health hazard in the Kingdom of Saudi Arabia owing to its sting, which has been known to cause cases of fatal anaphylactic shock (Dib et al., 1992, 1995). Al-Shahwan et al. (2006) reported a case of anaphylactic shock and since then several more such cases have been reported following samsum ant stings, some of which were really critical (Al-Anazi et al., 2009). Notwithstanding this negative reputation, however, B. sennaarensis can also be beneficial to humans: Dkhil et al. (2010) found that B. sennaarensis venom has an antiinflammatory effect that may be useful in the treatment of inflammatory skin diseases, whilst Badr et al. (2012) found that B. sennaarensis venom induces apoptosis in certain human breast cancer cells.
In this study, B. sennaarensis was detected in four provinces: ArRiyadh, Jazan, Najran and Eastern Province but was not detected in, Asir, Northern Frontiers, Tabouk, Makkah and Al-Madina. Collingwood (1985) suggested that the Arabian Peninsula is probably the northern limit of B. sennaarensis distribution. Different levels of occurrence of B. sennaarensis in the different regions of Saudi Arabia essentially confirm the non- indigenous status of the species. The population level also appears to depend on the geographical features of the location, with the high altitude of Asir, Makkah and Al-Madina regions, that each stands between 2000 and 3000 m above sea level, appearing to prohibit the occurrence of the species. Provinces such as Jazan and Najran are partially or completely Afrotropical in climate, since B. sennaarensis is indigenous to Africa this would explain their occurrence here. ArRiyadh and the Eastern region, meanwhile, are infested with a large number of ants due to large, frequent transport and heavy exchange of goods (Al-Khalifa et al., 2010). The Tabouk and Northern frontier regions, meanwhile, remain behind in terms of their developmental aspect and geosocial contacts, due to their distant location from the inhabited regions and have thus not yet been reached by these ants (Siddiqui and Al-Khalifa, 2013).
Yemen
Sharaf et al. (2017) - Brachyponera sennaarensis has invaded a wide range of habitats on Socotra, especially soil that is moist covered with the leaf litter of date palm trees. This species also commonly nests under rocks and objects associated with moist soils. Brachyponera sennaarensis has also invaded the relatively undisturbed valleys of the island where streams and denser vegetation are found. A nest was found under a stone under a dragon blood tree.
Association with Other Organisms
 Explore: Show all Associate data or Search these data. See also a list of all data tables or learn how data is managed.
Explore: Show all Associate data or Search these data. See also a list of all data tables or learn how data is managed.
Cestoda
- This species is a host for the cestode Raillietina tetragona (a parasite) in Sudan (Mohammed et al., 1988; Laciny, 2021).
Castes
Worker
,_Fig._S4.jpg)
{kind=link}
Images from AntWeb
     
| |
| Worker. Specimen code casent0048589. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by CAS, San Francisco, CA, USA. |
Queen
    
| |
| . | |
Male
    
| |
| . | |
Phylogeny
| Brachyponera |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Based on Chen et al., 2025. Note only selected species included.
Nomenclature
The following information is derived from Barry Bolton's Online Catalogue of the Ants of the World.
- sennaarensis. Ponera sennaarensis Mayr, 1862: 721 (w.) SUDAN.
- Type-material: holotype (?) worker.
- [Note: no indication of number of specimens is given.]
- Type-locality: Sudan (“Africa”): Sennaar (no collector’s name).
- Type-depository: NHMW.
- Santschi, 1910c: 350 (q.); Forel, 1910c: 245 (m.); Wheeler, G.C. & Wheeler, J. 1971b: 1207 (l.).
- Combination in Euponera (Brachyponera): Emery, 1901a: 47;
- combination in Pachycondyla: Brown, in Bolton, 1995b: 309;
- combination in Brachyponera: Wheeler, G.C. & Wheeler, J. 1971b: 1207; Schmidt, C.A. & Shattuck, 2014: 81.
- Status as species: Roger, 1863b: 16; Mayr, 1863: 450; Emery, 1877b: 366; Mayr, 1879: 662 (in key); Emery, 1881b: 528; André, 1884b: 538; Emery, 1892a: 111; Emery, 1892d: 557; Dalla Torre, 1893: 42; Emery, 1893f: 256; Forel, 1894b: 75; Mayr, 1895: 125; Emery, 1895a: 177; Emery, 1895f: 337; Emery 1897e: 597; Emery, 1899e: 474; Mayr, 1904b: 2; Forel, 1907a: 5; Forel, 1907g: 75; Forel, 1910c: 245; Santschi, 1910c: 350; Stitz, 1910: 130; Emery, 1911d: 84; Karavaiev, 1911: 3; Forel, 1913a: 108; Forel, 1913b: 307; Forel, 1913h: 347; Santschi, 1914b: 50; Santschi, 1914d: 318; Arnold, 1915: 72 (redescription); Stitz, 1916: 372; Wheeler, W.M. 1922a: 84, 777; Stitz, 1923: 144; Viehmeyer, 1923: 86; Menozzi, 1930b: 80; Menozzi, 1931a: 154; Santschi, 1933b: 96; Santschi, 1935b: 261; Santschi, 1937a: 49; Santschi, 1937d: 215; Finzi, 1939a: 154; Menozzi, 1939c: 99; Menozzi, 1942: 166; Weber, 1943c: 298; Eidmann, 1944: 469; Bernard, 1950b: 285; Menozzi & Consani, 1952: 60; Prins, 1963: 99; Collingwood, 1985: 240; Bolton, 1995b: 309; Collingwood & Agosti, 1996: 310; Collingwood, Tigar & Agosti, 1997: 506; Collingwood, et al. 2004: 478; Paknia, et al. 2008: 155; Collingwood, et al. 2011: 408; Hita Garcia, et al. 2013: 221; Wetterer, 2013a: 13; Borowiec, L. 2014: 141 (see note in bibliography); Bharti, Guénard, et al. 2016: 49; Sharaf, Fisher, et al. 2017: 51; Sharaf, Fisher, et al. 2018: 33; Madl, 2019: 16; Sharaf, Abdel-Dayem, et al. 2020: 554; Sharaf, Wetterer, et al. 2022: 86.
- Senior synonym of sorghi: Emery, 1899e: 474; Wheeler, W.M. 1922a: 777; Menozzi, 1930b: 80; Bolton, 1995b: 309.
- Distribution
- Afrotropical: Angola, Botswana, Burkina Faso, Burundi, Cameroon, Central African Republic, Chad, Congo, Democratic Republic of Congo, Equatorial Guinea, Eritrea, Ethiopia, Gabon, Gambia, Ghana, Guinea, Ivory Coast, Kenya, Liberia, Malawi, Mali, Mozambique, Niger, Nigeria, Senegal, Sierra Leone, Somalia, South Africa, South Sudan, Sudan, Tanzania, Uganda, Zambia, Zimbabwe.
- Oriental: India.
- Palaearctic: Bahrain, Cape Verde, Egypt, Iran, Kuwait, Oman, Qatar, Saudi Arabia, United Arab Emirates, Yemen.
- Current subspecies: nominal plus decolor, ruginota.
- sorghi. Ponera sorghi Roger, 1863a: 169 (w.) SUDAN.
- Type-material: holotype (?) worker.
- [Note: no indication of number of specimens is given.]
- Type-locality: Sudan: “Afrika, vom weissen fluss” (= White Nile), “in den Saaman von Sorghum Durra” (no collector’s name) (received from Gredler).
- [Note: Mayr, 1863: 450, merely says “Afrika”; Emery, 1911d: 84, has “Sudan”; Wheeler, W.M. 1922a: 777, has “Anglo-Egyptian Sudan: Sennar”.
- Type-depository: MNHU.
- Combination in Euponera (Brachyponera): Emery, 1911d: 84.
- Status as species: Roger, 1863b: 16; Mayr, 1863: 450; Dalla Torre, 1893: 42; Emery, 1911d: 84; Donisthorpe, 1942a: 27.
- Junior synonym of sennaarensis: Emery, 1899e: 474; Wheeler, W.M. 1922a: 777; Menozzi, 1930b: 80; Bolton, 1995b: 309.
Description
References
- Abdel-Dayem, M., Al Dhafer, H., Aldawood, A., Sharaf, M. 2021. An update to the taxonomy and distribution of the Arabian Tapinoma Foerster, 1850 (Hymenoptera: Formicidae) with an illustrated key and remarks on habitats. Biodiversity Data Journal 9, e66058 (doi:10.3897/bdj.9.e66058).
- Al-Khalifa, M.S., Ahmed, A.M., Mashaly, A.M.A., Al-Mekhalfi, F.A., Khaiil, G., Siddiqui, M.I., Ali, M.F. 2010. Studies on the distribution of Pachycondyla sennaarensis (Hymenoptera: Formicidae: Ponerinae) in Saudi Arabia. 1. Ar-Riyadh region. Pakistan Journal of Zoology, 42: 707-713.
- Al-Khalifa, M.S., Mashaly, A.M.A., Siddiqui, M.I., Al-Mekhlafi, F.A. 2015. Samsum ant, Brachyponera sennaarensis (Formicidae: Ponerinae): Distribution and abundance in Saudi Arabia. Saudi Journal of Biological Sciences 22, 575–579.
- Arnold, G. 1915. A monograph of the Formicidae of South Africa. Part I. Ponerinae, Dorylinae. Ann. S. Afr. Mus. 14: 1-159 (page 73, see also)
- Ashigar, M.A., Ab Majid, A.H. 2020. Diversity, abundance, and foraging behavior of ants (Hymenoptera: Formicidae) scavenging on American Cockroach in various habitats of Nasarawa State, Nigeria. Pertanika Journal of Tropical Agricultural Science 43: 503-521 (doi:10.47836/pjtas.43.4.07).
- Billen and Al-Khalifa. 2018. Morphology and ultrastructure of the venom gland in the ant Brachyponera sennaarensis. Asian Myrmecology. 10:e010005:1-9. doi:10.20362/am.010005
- Billen, J. and M. S. Al-Khalifa. 2015. Morphology and ultrastructure of the pro- and postpharyngeal glands in workers of Brachyponera sennaarensis. Sociobiology. 62:270-275. doi:10.13102/sociobiology.v62i2.270-275
- Billen, J., Al-Khalifa, M.S., Silva, R.R. 2017. Pretarsus structure in relation to climbing ability in the ants Brachyponera sennaarensis and Daceton armigerum. Saudi Journal of Biological Sciences 24, 830–836 (doi:10.1016/j.sjbs.2016.06.007).
- Billen, J., Ito, F. 2018. Novel thoracic glands in the ant Myopias hollandi. Arthropod Structure, Development 47, 229–237 (doi:10.1016/J.ASD.2018.04.005).
- Billen, J., Khalife, A., Ito, F., Anh, N.D., Esteves, F.A. 2021. The basitarsal sulcus gland, a novel exocrine structure in ants. Arthropod Structure, Development 61, 101041 (doi:10.1016/j.asd.2021.101041).
- Billen, J.P.J. 2019. Diversidad y morfología de las glándulas exocrinas en las hormigas. Pp. 165-174 in: Fernández, F., Guerrero, R.J., Delsinne, T. (eds.) 2019d. Hormigas de Colombia. Bogotá: Universidad Nacional de Colombia, 1198 pp.
- Borowiec, L. 2014. Catalogue of ants of Europe, the Mediterranean Basin and adjacent regions (Hymenoptera: Formicidae). Genus (Wroclaw) 25(1-2): 1-340.
- Borowiec, L., Salata, S. 2018. Notes on ants (Hymenoptera: Formicidae) from Gambia (Western Africa). Annals of the Upper Silesian Museum in Bytom, Entomology 26 (online 010), 1-13 (doi:10.5281/ZENODO.1243767).
- Brown, W. L., Jr. 1995a. [Untitled. Taxonomic changes in Pachycondyla attributed to Brown.] Pp. 302-311 in: Bolton, B. A new general catalogue of the ants of the world. Cambridge, Mass.: Harvard University Press, 504 pp. (page 309, combination in Pachycondyla)
- Chen, C., Yu, Y., Yi, C. 2025. Revision of the Chinese species of the genus Brachyponera Emery, 1900 (Hymenoptera, Formicidae), with a key to the world species of the genus. ZooKeys 1230: 247-286. (doi:10.3897/zookeys.1230.140159).
- Collingwood, C. A., Pohl, H., Guesten, R., Wranik, W. and van Harten, A. 2004. The ants (Insecta: Hymenoptera: Formicidae) of the Socotra Archipelago. Fauna of Arabia. 20:473-495.
- Collingwood, C.A., Agosti, D., Sharaf, M.R., van Harten, A. 2011. Order Hymenoptera, family Formicidae. Arthropod fauna of the UAE 4: 405-474.
- da Conceiao, E. S., J. H. C. Delabie, T. M. C. Della Lucia, A. D. Costa-Neto, and J. D. Majer. 2015. Structural changes in arboreal ant assemblages (Hymenoptera: Formicidae) in an age sequence of cocoa plantations in the south-east of Bahia, Brazil. Austral Entomology. 54:315-324. doi:10.1111/aen.12128
- Dejean A & Lachaud J-P 1994. Ecology and behavior of the seed-eating ponerine ant Brachyponera sennaarensis. Insectes Soc. 41: 191-210.
- Egbon, I.N., Osabuohien, I.P. 2022. First checklist, species richness and diversity of leaf-litter dwelling ants (Hymenoptera: Formicidae) in ancient Benin moat, Nigeria. Animal Research International 19(3): 4634–4642.
- Emery, C. 1901b. Notes sur les sous-familles des Dorylines et Ponérines (Famille des Formicides). Ann. Soc. Entomol. Belg. 45: 32-54 (page 47, Combination in Euponera (Brachyponera))
- Esteves, F.A., Fisher, B.L. 2021. Corrieopone nouragues gen. nov., sp. nov., a new Ponerinae from French Guiana (Hymenoptera, Formicidae). ZooKeys 1074, 83–173 (doi:10.3897/zookeys.1074.75551).
- Forel, A. 1910c. Ameisen aus der Kolonie Erythräa. Gesammelt von Prof. Dr. K. Escherich (nebst einigen in West-Abessinien von Herrn A. Ilg gesammelten Ameisen). Zool. Jahrb. Abt. Syst. Geogr. Biol. Tiere 29: 243-274 (page 245, male described)
- Lachaud, J.-P., Déjean, A. 1994. Predatory behavior of a seed-eating ant: Brachyponera senaarensis. Entomologia Experimentalis et Applicata 72(2), 145–155 (doi:10.1111/j.1570-7458.1994.tb01812.x).
- Mayr, G. 1862. Myrmecologische Studien. Verh. K-K. Zool.-Bot. Ges. Wien 12: 649-776 (page 721, worker described)
- Menozzi, C. 1930b. Formiche della Somalia italiana meridionale. Memorie della Societa Entomologica Italiana 9: 76-130 (page 80, Senior synonym of sorghi)
- Mohammed, O.B., Hussein, H.S., Elowni, E.E. 1988. The ant, Pachycondyla sennaarensis (Mayr) as an intermediate host for the poultry cestode, Raillietina tetragona (Molin). Veterinary Research Communications 12: 325-327 (doi:10.1007/BF00343251).
- Pashaei Rad, S., Taylor, B., Torabi, R., Aram, E., Abolfathi, G., Afshari, R., Borjali, F., Ghatei, M., Hediary, F., Jazini, F., Heidary Kiah, V., Mahmoudi, Z., Safariyan, F., Seiri, M. 2018. Further records of ants (Hymenoptera: Formicidae) from Iran. Zoology in the Middle East 64, 145-159 (doi:10.1080/09397140.2018.1442301).
- Richter, A., Hita Garcia, F., Keller, R.A., Billen, J., Economo, E.P., Beutel, R.G. 2020. Comparative analysis of worker head anatomy of Formica and Brachyponera (Hymenoptera: Formicidae). Senckenberg Gesellschaft für Naturforschung 78(1), 133–170 (doi:10.26049/ASP78-1-2020-06).
- Santschi, F. 1910c [1909]. Formicides nouveaux ou peu connus du Congo français. Ann. Soc. Entomol. Fr. 78: 349-400 (page 350, queen described)
- Schmidt, C.A. & Shattuck, S.O. 2014. The higher classification of the ant subfamily Ponerinae (Hymenoptera: Formicidae), with a review of ponerine ecology and behavior. Zootaxa 3817, 1–242 (doi:10.11646/zootaxa.3817.1.1).
- Sharaf, M. R. , B. L. Fisher, H. M. Al Dhafer, A. Polaszek and A. S. Aldawood. 2018. Additions to the ant fauna (Hymenoptera: Formicidae) of Oman: an updated list, new records and a description of two new species. Asian Myrmecology. 9:e010004; 1-38. doi:10.20362/am.010004
- Sharaf, M. R., Wetterer, J. K., Mohamed, A. A., Aldawood, A. S. 2022. Faunal composition, diversity, and distribution of ants (Hymenoptera: Formicidae) of Dhofar Governorate, Oman, with updated list of the Omani species and remarks on zoogeography. European Journal of Taxonomy 838: 1-106 (doi:10.5852/ejt.2022.838.1925).
- Sharaf, M.R., Abdel-Dayem, M.S., Mohamed, A.A., Fisher, B.L., Aldawood, A.S. 2020. A preliminary synopsis of the ant fauna (Hymenoptera: Formicidae) of Qatar with remarks on the zoogeography. Annales Zoologici 70: 533-560 (doi:10.3161/00034541anz2020.70.4.005).
- Sharaf, M.R., Fisher, B.L., Collingwood, C.A., Aldawood, A.S. 2017. Ant fauna (Hymenoptera: Formicidae) of the Socotra Archipelago (Yemen): zoogeography, distribution and description of a new species. Journal of Natural History 51, 317–378 (DOI 10.1080/00222933.2016.1271157).
- Siddiqui, M.I., Mashaly, A.M.A., Ahmed, A.M., Al-Mekhlafi, F.A., Al-Khalifa, M.S. 2010. Ultrastructure of antennal sensillae of the samsum ant, Pachycondyla sennaarensis (Hymenoptera: Formicidae). African Journal of Biotechnology, 9: 6956-6962.
- Taylor, B., Agoinon, N., Sinzogan, A., Adandonon, A., Kouaguou, Y. N., Bello, S., Wargui, R., Anato, F., Ouagoussounon, I., Houngbo, H., Tchibozo, S., Todjihounde, R., Vayssieres, J.F. 2018. Records of ants (Hymenoptera: Formicidae) from the Republic of Benin, with particular reference to the mango farm ecosystem. Journal of Insect Biodiversity 8(1): 6-29 (doi:10.12976/jib/2018.08.1.2).
- Tirgari, S.; Paknia, O. 2005. First record of the ponerine ant Pachycondyla sennaarensis (Hymenoptera: Formicidae) in Iran, with notes on its ecology. Zoology in the Middle East 34:67-70.
- Wetterer, J.K. 2013. Geographic spread of the samsum or sword ant, Pachycondyla (Brachyponera) sennaarensis (Hymenoptera: Formicidae). Myrmecological News 18, 13-18.
- Wheeler, G. C.; Wheeler, J. 1971b. Ant larvae of the subfamily Ponerinae: second supplement. Ann. Entomol. Soc. Am. 6 64: 1197-1217 (page 1207, larva described, Combination in Brachyponera)
- Wheeler, W. M. 1922j. Ants of the American Museum Congo expedition. A contribution to the myrmecology of Africa. VIII. A synonymic list of the ants of the Ethiopian region. Bull. Am. Mus. Nat. Hist. 45: 711-1004 (page 777, Senior synonym of sorghi)
- Xu, W., He, H., Billen, J. 2020. Morphology of the exocrine glands associated with the maxillolabial complex in the ant Camponotus japonicus Mayr, 1866 (Hymenoptera: Formicidae). Insectes Sociaux. (doi:10.1007/s00040-020-00793-2).
References based on Global Ant Biodiversity Informatics
- Borowiec L., and S. Salata. 2018. Notes on ants (Hymenoptera: Formicidae) from Gambia (Western Africa). Annals of the Upper Silesian Museum in Bytom Entomology 26: 1-13.
- Braet Y., and B. Taylor. 2008. Mission entomologique au Parc National de Pongara (Gabon). Bilan des Formicidae (Hymenoptera) recoltes. Bulletin S. R. B. E./K.B.V.E. 144: 157-169.
- Collingwood C.A., and A. Van Harten. 1993. The ants (Hymenoptera: Formicidae) of the Cape Verde Islands. Beitrag zur Fauna und Flora der Kapverdischen Inseln : Ergebnisse des 5. Symposiums in Leiden 159: 411-414.
- Diame L., B. Taylor, R. Blatrix, J. F. Vayssieres, J. Y. Rey, I. Grechi, and K. Diarra. 2017. A preliminary checklist of the ant (Hymenoptera, Formicidae) fauna of Senegal. Journal of Insect Biodiversity 5(15): 1-16.
- Dieng M. M., A. B. Ndiaye, C. T. Ba, and B. Taylor. 2016. Les fourmis (Hymenoptera, Formicidae) de l’enclos d’acclimatation de Katane de la reserve de faune du Ferlo nord (Senegal). Int. J. Biol. Chem. Sci. 10(4): 1626-1636.
- Donisthorpe H. 1942. The Formicidae (Hym.) of the Armstrong College Expedition to the Siwa Oasis. Annals and Magazine of Natural History (11)9: 26-33.
- Emery C. 1911. Hymenoptera. Fam. Formicidae. Subfam. Ponerinae. Genera Insectorum 118: 1-125.
- Emery C. 1915. Formiche raccolte nell'Eritrea dal Prof. F. Silvestri. Bollettino del Laboratorio di Zoologia Generale e Agraria della Reale Scuola Superiore d'Agricoltura. Portici 10: 3-26.
- Emery, C. "Catalogo delle formiche esistenti nelle collezioni del Museo Civico di Genova. Parte prima. Formiche provenienti dal Viaggio dei signori Antinori, Beccari e Issel nel Mar Rosso e nel paese dei Bogos. [concl.]." Annali del Museo Civico di Storia Naturale 9 (1877): 363-381.
- Finzi B. 1939. Materiali zoologici dell'Eritrea raccolti da G. Müller durante la spedizione dell'Istituto Sieroterapico Milanese e conservati al Museo di Trieste. Parte III. Hymenoptera: Formicidae. Atti del Museo Civico di Storia Naturale di Trieste 14: 153-168.
- Forel A. 1907. Ameisen von Madagaskar, den Comoren und Ostafrika. Wissenschaftliche Ergebnisse. Reise in Ostafrika 2: 75-92.
- Forel A. 1910. Ameisen aus der Kolonie Erythräa. Gesammelt von Prof. Dr. K. Escherich (nebst einigen in West-Abessinien von Herrn A. Ilg gesammelten Ameisen). Zoologische Jahrbücher. Abteilung für Systematik, Geographie und Biologie der Tiere 29: 243-274.
- Garcia F.H., Wiesel E. and Fischer G. 2013.The Ants of Kenya (Hymenoptera: Formicidae)Faunal Overview, First Species Checklist, Bibliography, Accounts for All Genera, and Discussion on Taxonomy and Zoogeography. Journal of East African Natural History, 101(2): 127-222
- IZIKO South Africa Museum Collection
- Karavaiev V. 1911. Ameisen aus Aegypten und dem Sudan. Rus. Entomol. Obozr. 11: 1-12.
- Kouakou L. M. 2015. Evaluation de la diversite des especes de fourmis anthropophiles, natives, exotiques et potentielles invasives en Cote d'Ivoire. Royal Belgian Institute of Natural Sciences
- Kouakou L. M. M., K. Yeo, K. Ouattara, W. Dekoninck, T. Delsinne, and S. Konate. 2018. Investigating urban ant community (Hymenoptera: Formicidae) in port cities and in major towns along the border in Côte d’Ivoire: a rapid assessment to detect potential introduced invasive ant species. Journal of Animal and Plant Sciences 36(1): 5793-5811.
- Kouakou L. M. M., W. Dekoninck, M. Kone, T. Delsinne, K. Yeo, K. Ouattara, and S. Konate. 2018. Diversity and distribution of introduced and potentially invasive ant species from the three main ecoregions of Côte d’Ivoire (West Africa). Belgian Journal of Zoology 148 (1): 83–103.
- Lévieux J., and T. Diomande. 1978. La nutrition des fourmis granivores. II. Cycle d'activite et regime alimentaire de Brachyponera senaarensis (Mayr) (Hymenoptera, Formicidae). Insectes Sociaux 25(3): 187-196.
- Lévieux J. 1972. Les fourmis de la savane de Lamto (Côte d'Ivoire): éléments de taxonomie. Bulletin de l'Institut Fondamental d'Afrique Noire. Série A. Sciences Naturelles 34: 611-654.
- Madl M. 2019. Notes on the ant fauna of Eritrea (Insecta: Hymenoptera: Formicidae): type specimens deposited in the Natural History Museum Vienna (Austria) and a preliminary checklist. Ann. Naturhist. Mus. Wien, B 121: 9-18.
- Medler J. T. 1980: Insects of Nigeria - Check list and bibliography. Mem. Amer. Ent. Inst. 30: i-vii, 1-919.
- Menozzi C. 1930. Formiche della Somalia italiana meridionale. Memorie della Società Entomologica Italiana. 9: 76-130.
- Menozzi C. 1939. Hymenoptera Formicidae. Missione Biologica nel Paese dei Borana. 3: 97-110.
- Menozzi C. 1942. Formiche dell'isola Fernando Poo e del territorio del Rio Muni (Guinea Spagnola). 24. Beitrag zu den wissenschaftlichen Ergebnissen der Forschungsreise H. Eidmann nach Spanisch-Guinea 1939 bis 1940. Zoologischer Anzeiger 140: 164-182.
- Menozzi C., M. Consani. 1952. Missione biologica Sagan-Omo diretta dal Prof. E. Zavattari. Hymenoptera Formicidae. Rivista di Biologia Coloniale 11: 57-71.
- Prins A. J. 1963. A list of the ants collected in the Kruger National Park with notes on their distribution. Koedoe 6: 91-108.
- Prins A. J. 1964. Revised list of the ants collected in the Kruger National Park. Koedoe 7: 77-93.
- Rafinejad J., A. Zareii, K. Akbarzadeh, M. Azad, F. Biglaryan, S. Doosti, and M. M. Sedaghat. 2009. Faunestic study of ants with emphasis on the health risk of stinging ants in Qeshm Island, Iran. Iranian J. Arthropod-Borne Dis 3(1): 53-59.
- Santschi F. 1910. Formicides nouveaux ou peu connus du Congo français. Annales de la Société Entomologique de France 78: 349-400.
- Santschi F. 1914. Formicides de l'Afrique occidentale et australe du voyage de Mr. le Professeur F. Silvestri. Bollettino del Laboratorio di Zoologia Generale e Agraria della Reale Scuola Superiore d'Agricoltura. Portici 8: 309-385.
- Santschi F. 1935. Hymenoptera. I. Formicidae. Mission Scientifique de l'Omo 2: 255-277.
- Santschi F. 1937. Résultats de la Mission scientifique suisse en Angola (2me voyage) 1932-1933. Fourmis angolaises. Revue Suisse de Zoologie. 44: 211-250.
- Stitz H. 1910. Westafrikanische Ameisen. I. Mitteilungen aus dem Zoologischen Museum in Berlin 5: 125-151.
- Stitz H. 1916. Formiciden. Ergebnisse der Zweiten Deutschen Zentral-Afrika Expedition 1: 369-405.
- Taylor B. 1976. Ants of the Nigerian Forest Zone (Hymenoptera: Formicidae). I. Ponerinae, Cerapachyinae, Pseudomyrmecinae. Cocoa Research Institute of Nigeria Technical Bulletin Series 4: 1-41.
- Taylor B., N. Agoinon, A. Sinzogan, A. Adandonon, Y. N'Da Kouagou, S. Bello, R. Wargui, F. Anato, I. Ouagoussounon, H. Houngbo, S. Tchibozo, R. Todjhounde, and J. F. Vayssieres. 2018. Records of ants (Hymenoptera: Formicidae) from the Republic of Benin, with particular reference to the mango farm ecosystem. Journal of Insect Biodiversity 8(1): 006–029.
- Viehmeyer H. 1923. Wissenschaftliche Ergebnisse der mit Unterstützung der Akademie der Wissenschaften in Wien aus der Erbschaft Treitl von F. Werner unternommenen zoologischen Expedition nach dem anglo-ägyptischen Sudan (Kordofan) 1914. VII. Hymenoptera A. Formicidae. Denkschriften der Akademie der Wissenschaften in Wien. Mathematisch-Naturwissenschaftliche Klasse 98: 83-94.
- Weber N. A. 1943. The ants of the Imatong Mountains, Anglo-Egyptian Sudan. Bulletin of the Museum of Comparative Zoology 93: 263-389.
- Wetterer J. K. 2013. Geographic spread of the samsum or sword ant, Pachycondyla (Brachyponera) sennaarensis (Hymenoptera: Formicidae). Myrmecological News 18: 13-18.
- Wheeler W. M. 1922. Ants of the American Museum Congo expedition. A contribution to the myrmecology of Africa. II. The ants collected by the American Museum Congo Expedition. Bulletin of the American Museum of Natural History 45: 39-269.
- Wheeler W. M. 1922. Ants of the American Museum Congo expedition. A contribution to the myrmecology of Africa. VIII. A synonymic list of the ants of the Ethiopian region. Bulletin of the American Museum of Natural History 45: 711-1004
- Pages using DynamicPageList3 parser function
- Facultatively polygynous
- Tandem running
- Invasive
- North temperate
- North subtropical
- Tropical
- South subtropical
- Photo Gallery
- Cestode Associate
- Host of Raillietina tetragona
- Species
- Extant species
- Formicidae
- Ponerinae
- Ponerini
- Brachyponera
- Brachyponera sennaarensis
- Ponerinae species
- Ponerini species
- Brachyponera species