Formica aquilonia
| Formica aquilonia | |
|---|---|

| |
| Conservation status | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Arthropoda |
| Class: | Insecta |
| Order: | Hymenoptera |
| Family: | Formicidae |
| Subfamily: | Formicinae |
| Tribe: | Formicini |
| Genus: | Formica |
| Species: | F. aquilonia |
| Binomial name | |
| Formica aquilonia Yarrow, 1955
| |
| Synonyms | |
| |



One of a number of Palearctic wood ants that build complexes of large mound nests. Gilev et al. (2015): A widespread and common species of red wood ant that inhabits the entire taiga belt of Eurasia (Dlussky, 1967). Similar to all red wood ants, this species displays a high degree of morphological variation.
| At a Glance | • Polygynous • Temporary parasite • Supercolonies • Diploid male |
Photo Gallery

Identification
Collingwood (1979) - Bicoloured with dark markings on head and promesonotum varying in size and intensity - generally not as brightly coloured or as large as Formica rufa. In the typical form distinct outstanding hairs fringe the posterior border of the head but do not occur forward towards the eyes as in Formica lugubris. In many samples from South Norway and South Finland these hairs may be hard to find or absent. Erect hairs on gula and dorsum of alitrunk variable, usually short and sparse. Eyes with distinct short hairs but much less prominent than in F. lugubris. Frons rather dull with close dense microsculpture, gaster very closely punctured. Suberect hairs on extensor surfaces of hind femora and tibiae always present but sometimes few. Antennal scapes bare. Head width of largest workers less than 2 mm. Length: 4.0-8.5 mm.
Keys including this Species
Distribution
Eastern Alps to Siberia, North Italy to North Norway (Collingwood 1979).
Latitudinal Distribution Pattern
Latitudinal Range: 70.377854° to 42.596228°.
| North Temperate |
North Subtropical |
Tropical | South Subtropical |
South Temperate |
- Source: AntMaps
Distribution based on Regional Taxon Lists
Palaearctic Region: Austria, Belarus, Bulgaria, China, Czechia, Democratic Peoples Republic of Korea, Denmark, Estonia, Finland, France, Georgia, Germany, Italy, Latvia, Lithuania, Mongolia, Montenegro, Norway, Poland, Romania, Russian Federation, Serbia, Slovenia, Sweden, Switzerland, Ukraine, United Kingdom of Great Britain and Northern Ireland (type locality).
Distribution based on AntMaps
Distribution based on AntWeb specimens
Check data from AntWeb
Countries Occupied
| Number of countries occupied by this species based on AntWiki Regional Taxon Lists. In general, fewer countries occupied indicates a narrower range, while more countries indicates a more widespread species. |
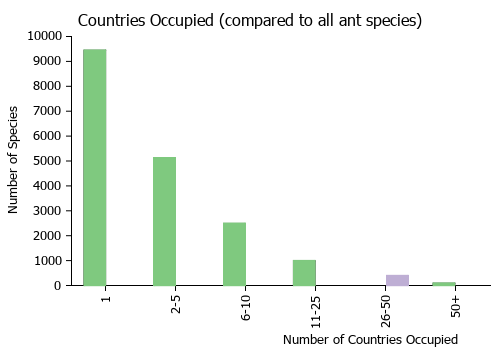
|
Biology
Gilev et al. (2015) - The results of studying the color variation of the red wood ant F. aquilonia in Northwestern Russia and Fennoscandia are presented. Pronounced phenotypic differentiation among the ant populations was observed, which may be interpreted in the light of postglacial dispersal of the species. Ants from Karelia and the environs of Moscow showed high similarity in their color variation. The share of a light morph (Pn3) increased in more northern localities, namely the Pasvik Reserve, and also in more western ones, namely St. Petersburg and Espoo. Thus, there were two vectors of ant dispersal: the northern and the western ones, coinciding with the known routes of postglacial recolonization by small mammals.

{kind=link}
Collingwood (1979) - This is undoubtedly the commonest wood ant in Fennoscandia. Large tracts of forest in the centre and north are dominated by this species which is usually found in large multicolonial groups with isolated nests being very rare. This is one of the least aggressive species of the F. rufa group. Long compact trails radiate from each nest to other nests or to aphid bearing trees and antagonism between neighbouring nests has not been observed. In South Norway and South Finland comparative hairlessness in many populations makes for confusion with the rather similar F. polyctena. Usually, however, if enough workers are collected a majority of at least 60 % of individuals will be found to have some projecting hairs at the back of the head.
However there is a form of this species found locally in the western suburbs of Helsinki to the Sjuntio district of Ab, within an area of about 40 km by 10 km, which is almost completely hairless in all parts of the body given as species specific by Yarrow (1955) for F. aquilonia. All castes moreover tend to be somewhat larger and more brightly coloured. This could well be a subspecies or species in the making. Its foraging habits have been studied by Rosengren (1971, 1977a, 1977b) under the name of Formica polyctena. This consistent degree of hairlessness has not been found elsewhere within the range of F. aquilonia, except perhaps in Esthonia, according to samples sent to H. Wuorenrinne by Professor V. Maavara.
The characteristics are as follows: only about 5 % or fewer workers in a series have an indication of short hairs projecting from the occipital comers of the head. Queens have no such hairs but occasional microscopic hairs have been detected on the basal face of the gaster in a very few specimens of about 50 examined. In the few males examined only 1 in 12 has projecting genal hairs. The reasons for retention as an infraspecific form of F. aquilonia include the close sculpturing of the worker frons, small eye hairs which are always present as in F. aquilonia while hairs on the extensor surface of the femora form a more or less close fringe as in F. aquilonia in a majority of the workers. The queen, which appears more brilliant than F. aquilonia, has extremely close micropunctures on the gaster as in that species (and in this character alone is quite unlike F. polyctena) while the male has fringing femoral hairs although specimens are also somewhat larger and more shining than F. aquilonia.
Novgorodova (2015) - Foragers and guards of Formica aquilonia are much more aggressive towards their topical competitors, the predatory ground beetles (Coleoptera, Carabidae), than towards mixo-phytophages (Dorosheva and Reznikova, 2006).
Foraging/Diet
Formica aquilonia collect large quantities of honeydew.
Novgorodova (2015b) investigated ant-aphid interactions of a dozen honeydew collecting ant species in Western Siberia pine and aspen-birch-pine forests (54°7´N, 83°06´E, 200 m, Novosibirsk) and mixed-grass-cereal steppes with aspen-birch groves (53°44´N, 78°02´E, 110 m, near Karasuk) in the Novosibirsk Region and coniferous forests in the northeastern Altai (north end of Lake Teletskoe, 51°48´N, 87°17´E, 434 m). All of the ants studied had workers that showed high fidelity to attending particular aphid colonies, i.e, individual ants tend to return to the same location, and group of aphids, every time they leave the nest. F. aquilonia's honeydew collecting activities were highly coordinated during the summer months when the aphids and ants were most active. Individual foragers specialized on specific tasks and could be classified as shepherds (collect honeydew), guards (protect aphids from competitors), scouts (search for new aphid colonies) and transporters (transport honeydew to the nest). Individuals performed the same type of work day after day, with groups of the same workers, thereby forming teams. In cooler months when aphids were still active foragers were less specialized: some ants were "on duty" (constantly present in a particular aphid colony, collecting honeydew and /or protecting aphids from various competitors) and others that showed no specialization. F. aquilonia tended: Symydobius oblongus (Heyden) and Chaitophorus populeti (Panzer).
Nesting Habits
Rybnikova and Kuznetsov (2015) studied nest complexes of wood ants in the Darwin Nature Reserve (Rybinsk Reservoir basin, Vologda and Yaroslavl Provinces, Russia). Their work assessed, in part, how wild boars and seasonal flooding may influence the survival and viability of wood ant colonies.
A complex of raised and transitional sphagnum bogs is developed in the central part of the peninsula. The better drained areas near the shores are occupied by a strip of upland forests from 1 to 5 km wide, mostly represented by green moss, tall moss, and complex pine forests blending into sphagnum pine forests closer to the bogs. Small patches of lichen pine forests are present in the raised areas. The biotic complex of the reserve is affected by the water level fluctuations in the reservoir, due to which its vast shallow peripheral areas are annually flooded and exposed. However, the water level not only changes seasonally within one year but also varies from year to year, so that high-water and low-water years occur. The destruction of mature ant nests by boars leads to complete elimination of many colonies and stimulates fragmentation of the surviving colonies in spring. The results of exogenous fragmentation of the damaged nests include a decrease in the number of large nests, loss of their growth potentials, depopulation, and degradation. Regular and largescale destruction of ant nests by boars leads to rapid degradation and dying off of whole nest complexes (Dyachenko, 1999; Efremov, 2013).
Observations of the ants have been carried out since 1997. The parameters recorded were the number of inhabited nests in the complexes (n), the basal diameter of the nest dome (d), and the diameter of the nest mound (D).
The complex of Kryukovaya Ridge (Formica aquilonia) included 54 nests. Since this locality was hard to access, the complex was completely examined only in 2010 and partly examined in 2013. Of the 54 nests, 42 revealed traces of heavy damage, up to total destruction; 40 nests (74.1%) were damaged by wild boars, one nest, by the bear Ursus arctos L., and one, by the badger Meles meles L. Most of the nests damaged by boars were almost completely destroyed, with only the circular mound remaining and a small central or peripheral dome built by the survivors. Most of the inhabited nests in this complex, namely 29 (53.7%), were located on the mounds of the previously destroyed larger nests. Under the conditions of total destruction, the diameters of the nest mound (D) and the dome base (d), i.e., the most objective indicators of the nest size dynamics, were considerably reduced. The mean diameter of the mound (D) of destroyed nests was 277 ± 71 cm (n = 29), whereas the mean diameter of the dome base (d) of inhabited nests was only 104 ± 45 cm (n = 25). Only 14 intact or weakly damaged nests were found in this complex, of which only two had the conical domes typical of active nests while 12 were flattened, i.e., not very viable. All these data indicated a considerable degree of degradation of the complex.
Association with Other Organisms
 Explore: Show all Associate data or Search these data. See also a list of all data tables or learn how data is managed.
Explore: Show all Associate data or Search these data. See also a list of all data tables or learn how data is managed.
- This species is a temporary parasite for the ant Formica fusca (a host) (Chernenko et al., 2013; de la Mora et al., 2021).
- This species is a temporary parasite for the ant Formica picea (a host) (Chernenko et al., 2013; de la Mora et al., 2021).
- This species is a xenobiont for the ant Formica fusca (a xenobiont) in Finland (Czechowski & Vepsalainen, 1999; Kanizsai et al., 2013) (Forest on rocks. Mound of F. lugubris).
- This species is a xenobiont for the ant Formicoxenus nitidulus (a xenobiont) (Holldobler & Wilson 1990; Busch 2001; Martin et al. 2007).
- This species is a xenobiont for the ant Lasius flavus (a xenobiont) in Finland (Czechowski, 2004; Kanizsai et al., 2013) (Different successional series of rocky habitats. Mound of Formica aquilonia.).
This species is associated with the aphids Aphis brohmeri, Aphis craccivora, Aphis evonymi, Aphis fabae, Aphis jacobaeae, Aphis pomi, Aphis subnitidae, Aphis viburni, Callipterinella tuberculata, Chaitophorus albitorosus, Chaitophorus albus, Chaitophorus populeti, Chaitophorus populialbae, Chaitophorus salicti, Cinara boerneri, Cinara laricis, Cinara piceae, Cinara piceicola, Cinara pinea, Cinara pini, Glyphina betulae, Rhopalosiphum padi, Schizaphis gramina, Schizaphis pyri and Symydobius oblongus (Saddiqui et al., 2019 and included references).
This species is a host for the following eulophid and braconid wasp parasites (Universal Chalcidoidea Database; Quevillon, 2018):
- Elasmosoma berolinense
- Neoneurus auctus
- Achrysocharoides niveipes
- Chrysocharis assis
- Chrysocharis assis
- Chrysocharis collaris
- Chrysocharis collaris
- Chrysocharis nephereus
- Chrysocharis nephereus
- Chrysocharis orbicularis
- Chrysocharis orbicularis
- Chrysocharis pilicoxa
- Chrysocharis pilicoxa
- Cirrospilus curvineurus
- Cirrospilus curvineurus
- Cirrospilus vittatus
- Cirrospilus vittatus
- Derostenus gemmeus
- Derostenus gemmeus
- Dicladocerus westwoodii
- Dicladocerus westwoodii
- Elachertus artaeus
- Elachterus artaeus
- Pediobius metallicus
- Pediobius metallicus
- Pnigalio longulus
- Pnigalio longulus
- Pnigalio pectinicornis
- Pnigalio pectinicornis
- Sympiesis gordius
- Sympiesis gordius
- Sympiesis sericeicornis
- Sympiesis sericeicornis
- Sympiesis xanthostoma
- Sympiesis xanthostoma
This species is a host for the beetle Monotoma angusticollis (Coleopotera: Monotomidae) (a myrmecophile) in Europe (Wagner et al., 2020).
The reported host/parasite relationship between Formica aquilonia and Formica cinerea should be investigated in the field as Chernenko et al. (2013) found 100% F. aquilonia queen mortality in lab introductions (de la Mora et al., 2021).
Flight Period
| X | X | ||||||||||
| Jan | Feb | Mar | Apr | May | Jun | Jul | Aug | Sep | Oct | Nov | Dec |
Source: antkeeping.info.
- Check details at Worldwide Ant Nuptial Flights Data, AntNupTracker and AntKeeping.
- Explore: Show all Flight Month data or Search these data. See also a list of all data tables or learn how data is managed.
Life History Traits
- Queen number: polygynous (Rissing and Pollock, 1988; Frumhoff & Ward, 1992)
- Queen mating frequency: multiple (Rissing and Pollock, 1988; Frumhoff & Ward, 1992)
- Colony type: supercolony
- Mean colony size: 400,000 (Zakharov, 1978; Beckers et al., 1989)
- Nest site: thatch mound
- Foraging behaviour: trunk trail (Zakharov, 1978; Beckers et al., 1989)
Castes
Worker
Images from AntWeb
   
| |
| Worker. Specimen code casent0172763. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by CAS, San Francisco, CA, USA. |
   
| |
| Worker. Specimen code casent0173151. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by CAS, San Francisco, CA, USA. |
Male
Diploid males are known to occur in this species (found in 1.2% of 334 examined nests) (Pamilo et al., 1994; Cournault & Aron, 2009).
Nomenclature
The following information is derived from Barry Bolton's Online Catalogue of the Ants of the World.
- schmidti. Formica rufa subsp. schmidti Ruzsky, 1926: 109 (w.) RUSSIA. [First available use of Formica rufa subsp. rufa var. schmidti Ruzsky, 1920: 78; unavailable name.] [Junior primary homonym of †schmidtii Heer, 1849: 138, above.] Synonymous with aquilonia: Dlussky, 1967a: 90, hence aquilonia the first available replacement name.
- aquilonia. Formica aquilonia Yarrow, 1955a: 31 (diagnosis in key), figs. 21, 25, 29, 33, 37, 41, 45, 49, 53, 57, 61, 65 (w.q.m.) GREAT BRITAIN. Synonymous with schmidti Ruzsky, 1920: 78 [Junior primary homonym of †schmidtii Heer, 1849: 138.]: Dlussky, 1967a: 90 and hence first available replacement name. See also: Kutter, 1977c: 271; Kupyanskaya, 1986a: 98; Gösswald, 1989: 19; Kupyanskaya, 1990: 197; Atanassov & Dlussky, 1992: 276.
Description
Karyotype
- See additional details at the Ant Chromosome Database.
- Explore: Show all Karyotype data or Search these data. See also a list of all data tables or learn how data is managed.
- n = 26 (Finland) (Rosengren et al., 1980).
References
- Alexandrovna, K.A. 2020. Red wood ants (Formica s. str.) as a method of biological protection in phytocenoses of the Mordovia Republic. In: Vavilov Readings - 2020: a collection of articles of the International Scientific and Practical Conference dedicated to the 100th anniversary of the discovery of the law of homological series and the 133rd anniversary of the birth of Academician NI Vavilov, November 24-25, 2020.
- Antonov, I.A., Bukin, Yu. S. 2016. Molecular phylogenetic analysis of the ant genus Formica L. (Hymenoptera: Formicidae) from Palearctic region. Russian Journal of Genetics, 52(8), 810–820 (doi:10.1134/s1022795416080020).
- Atanassov, N.; Dlussky, G. M. 1992. Fauna of Bulgaria. Hymenoptera, Formicidae. Fauna Bûlg. 22: 1-310 (page 276, see also)
- Badejo, O., Skaldina, O., Gilev, A., Sorvari, J. 2020. Benefits of insect colours: a review from social insect studies. Oecologia. (doi:10.1007/s00442-020-04738-1).
- Baer, B. 2011. The copulation biology of ants (Hymenoptera: Formicidae). Myrmecological News 14: 55-68.
- Baty, J.W., Bulgarella, M., Dobelmann, J., Felden, A., Lester, P.J. 2020. Viruses and their effects in ants (Hymenoptera: Formicidae). Myrmecological News 30: 213-228 (doi:10.25849/MYRMECOL.NEWS_030:213).
- Beckers R., Goss, S., Deneubourg, J.L., Pasteels, J.M. 1989. Colony size, communication and ant foraging Strategy. Psyche 96: 239-256 (doi:10.1155/1989/94279).
- Beresford, J. 2021. The role of hybrids in the process of speciation; a study of naturally occurring Formica wood ant hybrids. Academic Dissertation, University of Helsinki.
- Berkelhamer, R.C. 1980. Reproductive strategies in ants: A comparison of single-queened versus multiple-queened species in the subfamily Dolichoderinae (Hymenoptera: Formicidae). Ph.D. thesis, University of California, Berkeley.
- Bernasconi, C., Cherix, D., Seifert, B., Pamilo, P. 2011. Molecular taxonomy of the Formica rufa group (red wood ants) (Hymenoptera: Formicidae): a new cryptic species in the Swiss Alps? Myrmecological News 14: 37-47.
- Borowiec, L. 2014. Catalogue of ants of Europe, the Mediterranean Basin and adjacent regions (Hymenoptera: Formicidae). Genus (Wroclaw) 25(1-2): 1-340.
- Bulter, I. 2020. Hybridization in ants. Ph.D. thesis, Rockefeller University.
- Cerda, X., Arnan, X., Retana, J. 2013. Is competition a significant hallmark of ant (Hymenoptera: Formicidae) ecology? Myrmecological News 18: 131-147.
- Chen, Y., Zhou, S. 2017. Phylogenetic relationships based on DNA barcoding among 16 species of the ant genus Formica (Hymenoptera: Formicidae) from China. Journal of Insect Science 17(6): 117; 1–7 (doi:10.1093/jisesa/iex092).
- Chernenko, A., Vidal‐Garcia, M., Helantera, H., Sundstrom, L. 2013. Colony take‐over and brood survival in temporary social parasites of the ant genus Formica. Behavioral Ecology and Sociobiology 67: 727‐735 (doi:10.1007@s00265-013-1496-7).
- Cournault, L., Aron, S. 2009. Diploid males, diploid sperm production, and triploid females in the ant Tapinoma erraticum. Naturwissenschaften 96: 1393–1400 (doi:10.1007/s00114-009-0590-1).
- Csosz, S., Marko, B., Galle, L. 2011. The myrmecofauna (Hymenoptera: Formicidae) of Hungary: an updated checklist. North-Western Journal of Zoology 7: 55-62.
- Czechowski, W. 2004. Scarcity of sites suitable for nesting promotes plesiobiosis in ants (Hymenoptera: Formicidae). Entomologica Fennica 15(4). 211–218.
- Czechowski, W., Radchenko, A., Czechowska, W. 2002. The ants (Hymenoptera, Formicidae) of Poland. MIZ PAS Warsaw.
- de la Mora, A., Sankovitz, M., Purcell, J. 2020. Ants (Hymenoptera: Formicidae) as host and intruder: recent advances and future directions in the study of exploitative strategies. Myrmecological News 30: 53-71 (doi:10.25849/MYRMECOL.NEWS_030:053).
- Dekoninck, W., Maebe, K., Breyne, P., Hendrickx, F. 2014. Polygyny and strong genetic structuring within an isolated population of the wood ant Formica rufa. Journal of Hymenoptera Research 41, 95–111 (doi:10.3897/jhr.41.8191).
- Dlussky, G. M. 1967a. Ants of the genus Formica (Hymenoptera, Formicidae, g. Formica). Moskva: Nauka Publishing House, 236 pp. (page 90, Junior primary homonym of schmidtii Heer, 1850; and hence first available replacement name)
- Frizzi, F., Buonafede, L., Masoni, A., Balzani, P., Santini, G. 2022. Comparative analysis of facial coloration between introduced and source populations of the Red Wood Ant Formica paralugubris. Insects 13, 1137 (doi:10.3390/insects13121137).
- Frizzi, F., Masoni, A., Migliorini, M., Fanciulli, P.P., Cianferoni, F., Balzani, P., Giannotti, S., Davini, G., Frasconi Wendt, C., Santini, G. 2020. A comparative study of the fauna associated with nest mounds of native and introduced populations of the red wood ant Formica paralugubris. European Journal of Soil Biology 101, 103241. (doi:10.1016/j.ejsobi.2020.103241).
- Gilev, A. V., A. V. Mershchiev, and D. S. Malyshev. 2015. Interpretation of postglacial recolonization of Formica aquilonia (hymenoptera, formicidae) in Fennoscandia according to variability of workers' color. Zoologichesky Zhurnal. 94:1119-1124. doi:10.1134/S0013873815080011
- Goropashnaya, A.V., Fedorov, V.B., Seifert, B., Pamilo, P. 2012. Phylogenetic relationships of Palaearctic Formica species (Hymenoptera, Formicidae) based on mitochondrial Cytochrome b sequences. PLoS ONE 7, e41697 (doi:10.1371/journal.pone.0041697).
- Goropashnaya, Anna V.; Fedorov, Vadim B.; Pamilo, Pekka 2004. Recent speciation in the Formica rufa group ants (Hymenoptera, Formicidae): inference from mitochondrial DNA phylogeny. Molecular Phylogenetics and Evolution 32(1): 198-206 (mDNA sequence data)
- Gösswald, K. 1989. Die Waldameise. Band 1. Biologische Grundlagen, Ökologie und Verhalten. Wiesbaden: AULA-Verlag, xi + 660 pp. (page 19, see also)
- Helantera, H., Sundstrom, L. 2007. Worker reproduction in Formica ants. The American Naturalist 170: E15-E25.
- Jacobs, S. 2020. Population genetic and behavioral aspects of male mating monopolies in Cardiocondyla venustula (Ph.D. thesis).
- Kanizsai, O., Lőrinczi, G., Gallé, L. 2013. Nesting associations without interdependence: A preliminary review on plesiobiosis in ants. Psyche 2013, 238602 (doi:10.1155/2013/238602).
- Kupyanskaya, A. N. 1986b. Ants (Hymenoptera, Formicidae) from the northern part of the Far East. Pp. 91-102 in: Ler, P. A. (ed.) Systematics and ecology of insects from the Far East. Vladivostok: Akademiya Nauk SSSR, 155 pp. (page 98, see also)
- Kupyanskaya, A. N. 1990a. Ants of the Far Eastern USSR. Vladivostok: Akademiya Nauk SSSR, 258 pp. (page 197, see also)
- Kutter, H. 1977c. Hymenoptera, Formicidae. Insecta Helv. Fauna 6: 1-298.
- Nouhaud, P., Beresford, J., Kulmuni, J. 2022. Assembly of a hybrid Formica aquilonia × F. polyctena ant genome from a haploid male. Journal of Heredity 113(3), 353–359 (doi:10.1093/jhered/esac019).
- Nouhaud, P., Martin, S.H., Portinha, B., Sousa, V.C., Kulmuni, J. 2022. Rapid and predictable genome evolution across three hybrid ant populations. PLOS Biology, 2012, e3001914 (doi:10.1371/journal.pbio.3001914).
- Novgorodova, T. A. 2015b. Organization of honeydew collection by foragers of different species of ants (Hymenoptera: Formicidae): Effect of colony size and species specificity. European Journal of Entomology. 112:688-697. doi:10.14411/eje.2015.077
- Nygard, E. 2020. Thermal tolerance of two mound-building wood ants Formica aquilonia and Formica polyctena and their hybrids. Master's thesis, University of Helsinki.
- Pamilo, P., Kulmuni, J. 2022. Genetic identification of Formica rufa group species and their putative hybrids in northern Europe. Myrmecological News 32: 93-102 (doi:10.25849/MYRMECOL.NEWS_032:093).
- Pamilo, P., Sundström, L., Fortelius, W., Rosengren, R. 1994. Diploid males and colony-level selection in Formica ants. Ethology Ecology, Evolution 6, 221–235 (doi:10.1080/08927014.1994.9522996).
- Portinha, B., Avril, A., Bernasconi, C., Helanterä, H., Monaghan, J., Seifert, B., Sousa, V.C., Kulmuni, J., Nouhaud, P. 2021. Whole-genome analysis of multiple wood ant population pairs supports similar speciation histories, but different degrees of gene flow, across their European range. bioRxiv preprint (doi:10.1101/2021.03.10.434741).
- Punttila, P., Kilpeläinen, J. 2009. Distribution of mound-building ant species (Formica spp., Hymenoptera) in Finland: preliminary results of a national survey. Annales Zoologici Fennici 46: 1–15.
- Purkart, A., Repta, F. 2022. First record of Cryptopone ochracea (Mayr, 1855) (Hymenoptera: Formicidae) in Slovakia with notes on its nesting biology. Entomofauna carpathica 34(2): 85-94.
- Putyatina, T.S., Gilev, A.V., Grinkov, V.G., Markov, A.V. 2022. Variation in the colour pattern of the narrow-headed ant Formica exsecta (Hymenoptera: Formicidae) in European Russia. European Journal of Entomology 119, 58-68 (doi:10.14411/eje.2022.006).
- Radchenko, A.G., Fisher, B.L., Esteves, F.A., Martynova, E.V., Bazhenova, T.N., Lasarenko, S.N. 2023. Ant type specimens (Hymenoptera, Formicidae) in the collection of Volodymyr Opanasovych Karawajew. Communication 1. Dorylinae, Poneromorpha and Pseudomyrmecinae. Zootaxa, 5244(1), 1–32 (doi:10.11646/zootaxa.5244.1.1).
- Ruzsky, M. 1920. Ants of Kamchatka. Izv. Inst. Issled. Sib. 2: 76-80 (page 78, synonymous with schmidti)
- Rybnikova, I. A. and A. V. Kuznetsov. 2015. Complexes of Formica s. str. nests in the Darwin Nature Reserve and causes of their degradation. Entomological Review. 95:947-952. doi:10.1134/s0013873815080023
- Schifani, E. (2022). The new checklist of the Italian fauna: Formicidae. Biogeographia – The Journal of Integrative Biogeography 37, ucl006 (doi:10.21426/b637155803).
- Schultner, E., Pulliainen, U. 2020. Brood recognition and discrimination in ants. Insectes Sociaux 67, 11–34 (doi:10.1007/s00040-019-00747-3).
- Schultner, E., Saramaki, J., Helantera, H. 2016. Genetic structure of native ant supercolonies varies in space and time. Molecular Ecology 25, 6196–6213 (DOI 10.1111/mec.13912).
- Seifert, B. 2021. A taxonomic revision of the Palaearctic members of the Formica rufa group (Hymenoptera: Formicidae) – the famous mound-building red wood ants. Myrmecological News 31: 133-179 (doi:10.25849/MYRMECOL.NEWS_031:133).
- Siddiqui, J. A., Li, J., Zou, X., Bodlah, I., Huang, X. 2019. Meta-analysis of the global diversity and spatial patterns of aphid-ant mutualistic relationships. Applied Ecology and Environmental Research 17: 5471-5524 (doi:10.15666/aeer/1703_54715524).
- Siedlecki, I., Gorczak, M., Okrasińska, A., Wrzosek, M. 2021. Chance or necessity—The fungi co−occurring with Formica polyctena ants. Insects 12, 204 (doi:10.3390/insects12030204).
- Sundstrom, L. 1993. Genetic population structure and sociogenetic organisation in Formica truncorum (Hymenoptera; Formicidae). Behavioral Ecology and Sociobiology 33:345-354
- Wagner, G.K., Staniec, B., Zagaja, M., Pietrykowska-Tudruj, E. 2020. First insight into detailed morphology of monotomids, with comments on chaetotaxy and life history based on myrmecophilous Monotoma angusticollis. Bulletin of Insectology 73 (1): 11-27.
- Yarrow, I. H. H. 1955a. The British ants allied to Formica rufa L. (Hym., Formicidae). Trans. Soc. Br. Entomol. 12: 1-48 (page 31, figs. 21, 25, 29, 33, 37, 41, 45, 49, 53, 57, 61, 65 worker, queen, male described)
References based on Global Ant Biodiversity Informatics
- Antonov I. A. 2012. Ant complexes of Baikalsk town. The Bulletin of Irkutsk State University 4: 143-146.
- Antonov I. A. 2013. Ant Assemblages (Hymenoptera: Formicidae) of Cities of the Temperate Zone of Eurasia. Russian Journal of Ecology 44(6): 523526.
- ArtDatabanken Bugs (via GBIG)
- Banert P, and B. Pisarski. 1972. Mrówki (Formicidae) Sudetów. Fragmenta Faunistica (Warsaw) 18: 345-359.
- Baroni Urbani C., and C. A. Collingwood. 1976. A Numerical Analysis of the Distribution of British Formicidae (Hymenoptera, Aculeata). Verhandlungen der Naturforschenden Gesellschaft in Basel 85: 51-91.
- Baroni Urbani C., and C. A. Collingwood. 1977. The zoogeography of ants (Hymenoptera, Formicidae) in Northern Europe. Acta Zoologica Fennica 152: 1-34.
- Belskaya E., A. Gilev, and E. Belskii. 2017. Ant (Hymenoptera, Formicidae) diversity along a pollution gradient near the Middle Ural Copper Smelter, Russia. Environ Sci Pollut Res DOI 10.1007/s11356-017-8736-8
- Bernard F. 1967. Faune de l'Europe et du Bassin Méditerranéen. 3. Les fourmis (Hymenoptera Formicidae) d'Europe occidentale et septentrionale. Paris: Masson, 411 pp.
- Borowiec L. 2014. Catalogue of ants of Europe, the Mediterranean Basin and adjacent regions (Hymenoptera: Formicidae). Genus (Wroclaw) 25(1-2): 1-340.
- Buhl, J., K. Hicks, E.R. Miller, S. Persey, O. Alinvi and D.J.T. Sumpter. 2009. Shape and efficiency of wood ant foraging networks. Behavioral Ecology and Sociobiology 63:451-460
- Collingwood C. A. 1956. Ant hunting in France. Entomologist 89: 106-108.
- Collingwood C. A. 1961. Ants in the Scottish Highlands. Scotish Naturalist 70: 12-21.
- Collingwood C. A. 1963. Three ant species new to Norway. Entomologist's Record and Journal of Variation 75: 225-228.
- Collingwood C. A. 1971. A synopsis of the Formicidae of north Europe. Entomologist 104: 150-176
- Collingwood C.A. 1959. Ants in the Scottish Highlands. The Scottish Naturalist. 70: 12-21
- Collingwood C.A. 1959. Scandinavian Ants. Entomol. Rec. 71: 78-83
- Collingwood C.A. 1961. Ants in Finland. Entomol. Rec. 73: 190-195
- Collingwood, C. A. 1958b. A key to the species of ants (Hymenoptera, Formicidae) found in Britain. Trans. Soc. Br. Entomol. 13: 69-96
- Collingwood, C. A. 1964. The Identification of British Ants (Hym. Formicidae). Transactions of the Society for British Entomology. 16:93-121.
- Collingwood, C. A. 1974. A revised list of Norwegian ants (Hymenoptera: Formicidae). Norsk Entomologisk Tidsskrift 21: 31-35.
- Collingwood, C. A.. "The Formicidae (Hymenoptera) of Fennoscandia and Denmark." Fauna Entomologica Scandinavica 8 (1979): 1-174.
- Collingwood, C.A. 1958. A survey of Irish Formicidae. Proceedings of the Royal Irish Academy 59B:213-219
- Czechowski W., A. Radchenko, W. Czechowska and K. Vepsäläinen. 2012. The ants of Poland with reference to the myrmecofauna of Europe. Fauna Poloniae 4. Warsaw: Natura Optima Dux Foundation, 1-496 pp
- Dubovikoff D. A., and Z. M. Yusupov. 2018. Family Formicidae - Ants. In Belokobylskij S. A. and A. S. Lelej: Annotated catalogue of the Hymenoptera of Russia. Proceedingss of the Zoological Institute of the Russian Academy of Sciences 6: 197-210.
- Else G., B. Bolton, and G. Broad. 2016. Checklist of British and Irish Hymenoptera - aculeates (Apoidea, Chrysidoidea and Vespoidea). Biodiversity Data Journal 4: e8050. doi: 10.3897/BDJ.4.e8050
- Entomological Society of Latvia. 2003. http://leb.daba.lv/Formicidae.htm (Accessed on December 1st 2013).
- Field Museum Collection, Chicago, Illinois (C. Moreau)
- Gibb H., and T. Johansson. 2011. Field tests of interspecific competition in ant assemblages: revisiting the dominant red wood ants. Journal of Animal Ecology 80: 548-557.
- Gilev A. V., I. V. Kuzmin, V. A. Stolbov, and S. D. Sheikin. 2012. Materials on the fauna and ecology of ants (formicidae) Southern part of the Tyumen region. Tyumen State University Herald 6: 86-91.
- Glaser, F., M. J. Lush, and B. Seifert. "Rediscovered after 140 years at two localities: Myrmica myrmicoxena Forel, 1895 (Hymenoptera: Formicidae)." Myrmecological News 14 (2010): 107-111.
- Gratiashvili N., Barjadze S. 2008. Checklist of the ants (Formicidae Latreille, 1809) of Georgia. Proceedings of the Institute of Zoology (Tbilisi) 23: 130-146.
- Gulzar A. 2014. Classification and Distribution of ants in Kashgar, Xinjiang. Master's Thesis Shaanxi Normal University, 75 pages.
- Guénard B., and R. R. Dunn. 2012. A checklist of the ants of China. Zootaxa 3558: 1-77.
- Heatwole H. 1989. Changes in ant assemblages across an artic treeline. Revue d'Entomologie du Quebec 34(1-2): 10-22.
- Hågvar S. 2005. Altitudinal zonation of ants (Formicidae) in a steep fjord landscape in Sogndal, Western Norway. Norw. J. Entomol. 52: 3-12.
- Kipyatkov V. E. 1993. Annual cycles of development in ants: diversity, evolution, regulation. In: Proceedings of the Colloquia on Social Insects, vol. 2, 2d Colloquium, Rybnoye, 1992. Socium, St. Petersburg, pp 25-48.
- Kofler A. 1995. Nachtrag zur Ameisenfauna Osttirols (Tirol, Österreich) (Hymenoptera: Formicidae). Myrmecologische Nachrichten 1: 14-25.
- Kvamme T. 1982. Atlas of the Formicidae of Norway (Hymenoptera: Aculeata). Insecta Norvegiae 2: 1-56.
- Lelej A. S. 2012. Annotated catalogue of the Insects of Russian Far East. Volume 1. Hymenoptera. Dalnauka: Vladivostok. 635 p.
- Malozemova L. A. 1972. Ants of steppe forests, their distribution by habitats, and perspectives of their utilization for protection of forests (north Kazakhstan). [In Russian.]. Zoologicheskii Zhurnal 51: 57-68.
- Nielsen M. G. 2011. A check list of Danish ants and proposed common names. Ent. Meddr. 79: 13-18.
- Paukkunen J., and M. V. Kozlov. 2015. Stinging wasps, ants and bees (Hy menoptera: Aculeata) of the Murmansk region, Northwest Russia. — Entomol. Fennica. 26: 53–73.
- Pisarski B. 1969. Fourmis (Hymenoptera: Formicidae) de la Mongolie. Fragmenta Faunistica (Warsaw). 15: 221-236.
- Pisarski B. 1982. Ants (Hymenoptera: Formicoidea) of Warsaw and Mazovia. Memorabilia Zool. 36: 73-90.
- Punttila P., S. Koponen, and M. Saaristo. 1994. Colonisation of a burned mountain-birch forest by ants (Hymenoptera, Formicidae) in subarctic Finland. Polish Academy of Sciences, Museum and Institute of Zoology, Memorabilia Zoologica 48: 193-206.
- Punttila P., Y. Haila, J. Niemela, and T. Pajunen. 1994. Ant communities in fragments of old-growth taiga and managed surroundings. Ann. Zool. Fennici 31: 131-144.
- Punttila P., Y. Haila, and H. Tukia. 1996. Ant communities in taiga clearcuts: habitat effects and species interactions. Ecography 19: 16-28.
- Punttila P., and Y. Haila. 1996. Colonisation of a burned forest by ants in the southern Finnish Boreal forest. Silva Fennica 30(4): 421-435.
- Radchenko, A. 2005. Monographic revision of the ants (Hymenoptera, Formicidae) of North Korea. Annales Zoologici 55(2): 127-221.
- Ran H., and S. Y. Zhou. 2012. Checklist of chinese ants: formicomorph subfamilies (Hymenoptera: Formicidae) II. Journal of Guangxi Normal University: Natural Science Edition 30(4): 81-91.
- Ruzsky M. 1920. Ants of Kamchatka. Izv. Inst. Issled. Sib. 2: 76-80
- Seifert B. 1994. Die freilebenden Ameisenarten Deutschlands (Hymenoptera: Formicidae) und Angaben zu deren Taxonomie und Verbreitung. Abhandlungen und Berichte des Naturkundemuseums Görlitz 67(3): 1-44.
- Seifert B. 1996. Formica paralugubris nov. spec. - a sympatric sibling species of Formica lugubris from the western Alps (Insecta: Hymenoptera: Formicoidea: Formicidae). Reichenbachia 31: 193-201.
- Seifert B. 1998. Rote Liste der Ameisen. - in: M. Binot, R. Bless, P. Boye, H. Gruttke und P. Pretscher: Rote Liste gefährdeter Tiere Deutschlands. Bonn-Bad Godesberg 1998: 130-133.
- Seifert B. 2007. Die Ameisen Mittel- und Nordeuropas. Tauer: lutra Verlags- und Vertriebsgesellschaft, 368 pp.
- Sipura, M. 2002. Contrasting Effects of Ants on the Herbivory and Growth of Two Willow Species. Ecology 83(10):2680-2690
- Steiner F. M., S. Schödl, and B. C. Schlick-Steiner. 2002. Liste der Ameisen Österreichs (Hymenoptera: Formicidae), Stand Oktober 2002. Beiträge zur Entomofaunistik 3: 17-25.
- Suvak M. 2013. First record of Formica fennica Seifert, 2000 (Hymenoptera, Formicidae) in Norway. Norwegian Journal of Entomology 60: 73–80.
- Vele A., J. Holisa, J. Tragnerova. 2015. Ant succession on burned areas in forested landscape: a case study from the Bohemian Switzerland National Forest. Zpravy Lesnickeho Vyzkumu 60(1): 47-52.
- Vepsalainen K., H. Ikonene, and M. J. Koivula. 2008. The structure of ant assembalges in an urban area of Helsinki, southern Finland. Ann. Zool. Fennici 45: 109-127.
- Wu J. 1990. Taxonomic studies on the genus Formica of China (Hymenoptera: Formicidae). Forest Research 3: 1-8.
- Yarrow I. H. H. 1955. The British ants allied to Formica rufa L. (Hym., Formicidae). Trans. Soc. Br. Entomol. 12: 1-48.
- Zhuytszyuan D. 2016. The ants (Hymenoptera, Formicidae) Nizhne-Svirsky reserve and their environmental features. Master's thesis Saint Petersburg State University.
- Zryanin V. A., and T. A. Zryanina. 2007. New data on the ant fauna Hymenoptera, Formicidae in the middle Volga River Basin. Uspekhi Sovremennoi Biologii 127(2): 226-240.
- Pages using DynamicPageList3 parser function
- IUCN Red List near threatened species
- Polygynous
- Temporary parasite
- Supercolonies
- Diploid male
- Photo Gallery
- North temperate
- Nesting Notes
- Ant Associate
- Host of Formica fusca
- Host of Formica picea
- Host of Formicoxenus nitidulus
- Host of Lasius flavus
- Aphid Associate
- Host of Aphis brohmeri
- Host of Aphis craccivora
- Host of Aphis evonymi
- Host of Aphis fabae
- Host of Aphis jacobaeae
- Host of Aphis pomi
- Host of Aphis subnitidae
- Host of Aphis viburni
- Host of Callipterinella tuberculata
- Host of Chaitophorus albitorosus
- Host of Chaitophorus albus
- Host of Chaitophorus populeti
- Host of Chaitophorus populialbae
- Host of Chaitophorus salicti
- Host of Cinara boerneri
- Host of Cinara laricis
- Host of Cinara piceae
- Host of Cinara piceicola
- Host of Cinara pinea
- Host of Cinara pini
- Host of Glyphina betulae
- Host of Rhopalosiphum padi
- Host of Schizaphis gramina
- Host of Schizaphis pyri
- Host of Symydobius oblongus
- Braconid wasp Associate
- Host of Elasmosoma berolinense
- Host of Neoneurus auctus
- Eulophid wasp Associate
- Host of Achrysocharoides niveipes
- Host of Chrysocharis assis
- Host of Chrysocharis collaris
- Host of Chrysocharis nephereus
- Host of Chrysocharis orbicularis
- Host of Chrysocharis pilicoxa
- Host of Cirrospilus curvineurus
- Host of Cirrospilus vittatus
- Host of Derostenus gemmeus
- Host of Dicladocerus westwoodii
- Host of Elachertus artaeus
- Host of Elachterus artaeus
- Host of Pediobius metallicus
- Host of Pnigalio longulus
- Host of Pnigalio pectinicornis
- Host of Sympiesis gordius
- Host of Sympiesis sericeicornis
- Host of Sympiesis xanthostoma
- Beetle Associate
- Host of Monotoma angusticollis (Coleopotera: Monotomidae)
- FlightMonth
- Karyotype
- Species
- Extant species
- Formicidae
- Formicinae
- Formicini
- Formica
- Formica aquilonia
- Formicinae species
- Formicini species
- Formica species
- Need Body Text
- IUCN Red List