Atta colombica
| Atta colombica | |
|---|---|

| |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Arthropoda |
| Class: | Insecta |
| Order: | Hymenoptera |
| Family: | Formicidae |
| Subfamily: | Myrmicinae |
| Tribe: | Attini |
| Genus: | Atta |
| Species: | A. colombica |
| Binomial name | |
| Atta colombica Guérin-Méneville, 1844
| |
| Synonyms | |
| |


Photo Gallery

.jpg)
{kind=link}
Identification
Distribution
Latitudinal Distribution Pattern
Latitudinal Range: 14.624795° to -12.043°.
| North Temperate |
North Subtropical |
Tropical | South Subtropical |
South Temperate |
- Source: AntMaps
Distribution based on Regional Taxon Lists
Neotropical Region: Colombia (type locality), Costa Rica, Guatemala, Nicaragua, Panama, Peru.
Distribution based on AntMaps
Distribution based on AntWeb specimens
Check data from AntWeb
Countries Occupied
| Number of countries occupied by this species based on AntWiki Regional Taxon Lists. In general, fewer countries occupied indicates a narrower range, while more countries indicates a more widespread species. |

|
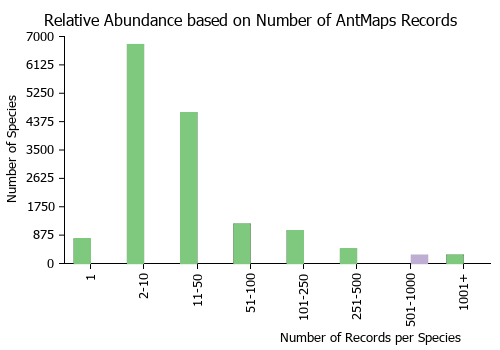
Estimated Abundance
| Relative abundance based on number of AntMaps records per species (this species within the purple bar). Fewer records (to the left) indicates a less abundant/encountered species while more records (to the right) indicates more abundant/encountered species. |
|
Biology
|
Dijkstra and Boomsma (2006) investigated the viability of worker produced eggs in Atta cephalotes, Atta sexdens and Atta colombica. Most Atta workers have rudimentary, non-functional ovaries in a queenright colony but a few, typically tending the queen, can produce trophic eggs (Dijkstra et al., 2005). These eggs are feed to the queen. It was not known if any worker eggs can produce males. Most worker eggs of A. colombica did not contain yolk but a few did and yielded morphologically correct but very small males. They hypothesize that worker reproduction in orphaned Atta field colonies is almost never successful because the last workers die before their sons can be raised to adulthood, but the importance of worker-laid trophic eggs for queen feeding has precluded the evolutionary loss of worker ovaries.
Foraging
Bochynek et al. (2017) studied foraging rhythms in a colony of A. colombica in Panama: Foraging on the control (leaf) trail was diurnal and occurred in discrete periods approximately 10 h long. Activity began at dawn, rose to a peak in early afternoon and declined toward evening, with no foraging at night, a typical pattern for Atta leaf foraging (Lewis et al., 1974a). Leaf foraging on the berry/leaf trail followed a similar pattern. The peak harvesting rates, about 100–150 laden ant returns per min., were similar to the mean rate of 108 laden returns per min. observed by Bruce and Burd (2012) for colonies of A. colombica and Atta cephalotes in Panama and Costa Rica. Thus, leaf foraging was typical of colonies at other times and places.
Fruit foraging was fundamentally different. Harvesting was continuous with no periods of inactivity. There was, nonetheless, a diel rhythm. Throughout most of a 24 h cycle, fruit carriers returned at approximately the same rate as leaf harvesters at peak activity, but activity slowed to about 50 returns per min. in early morning, at 07.00 hours, before returning to a plateau of about 100–150 returns per min. for the remainder of the cycle.
Association with Other Organisms
 Explore: Show all Associate data or Search these data. See also a list of all data tables or learn how data is managed.
Explore: Show all Associate data or Search these data. See also a list of all data tables or learn how data is managed.
- This species is a host for the phorid fly Apocephalus colombicus (a parasite) (phorid.net) (attacked).
- This species is a host for the phorid fly Apocephalus attophilus (a parasitoid) (Quevillon, 2018) (encounter mode primary; direct transmission; transmission outside nest).
- This species is a host for the phorid fly Eibesfeldtphora pala (a parasitoid) (Quevillon, 2018) (encounter mode primary; direct transmission; transmission outside nest).
- This species is a host for the fungus Ophiocordyceps subramanianii (a parasitoid) (Quevillon, 2018) (encounter mode primary; direct transmission; transmission outside nest).
Castes
Images from AntWeb
   
| |
| Syntype of Atta cephalotes erecta. Worker. Specimen code casent0912490. Photographer Z. Lieberman, uploaded by California Academy of Sciences. | Owned by NHMB, Basel, Switzerland. |
   
| |
| Syntype of Atta colombica tonsipes. Worker. Specimen code casent0912491. Photographer Z. Lieberman, uploaded by California Academy of Sciences. | Owned by NHMB, Basel, Switzerland. |
Phylogeny
| Atta |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Based on Barrera, C.A. et al., 2021. Note that only selected species are included.
Nomenclature
The following information is derived from Barry Bolton's Online Catalogue of the Ants of the World.
- colombica. Atta colombica Guérin-Méneville, 1844a: 422 (w.) COLOMBIA.
- Type-material: holotype (?) worker.
- [Note: no indication of number of specimens is given.]
- Type-locality: Colombia: (no further data).
- [Notes (i): Borgmeier, 1959b: 346, cites Colombia: Santa Marta as type-locality; (ii) according to Horn & Kahle, 1935: 99, Guérin’s Hymenoptera material was split between MNHN, MSNG, and ZSBS, but some are also present in NNML.]
- Type-depository: MNHN.
- [Misspelled as columbica by Roger, 1863b: 35, Mayr, 1865: 81, Forel, 1885a: 362, Emery, 1890b: 55, Dalla Torre, 1893: 152, Forel, 1899c: 33, Forel, 1912e: 179, Emery, 1913b: 259, Wheeler, W.M. 1923c: 5, Emery, 1924d: 353, Menozzi, 1927c: 268, Santschi, 1929f: 92, Weber, 1938b: 205, Borgmeier, 1939: 422, Bolton, 1995b: 75, and others.]
- Forel, 1913l: 239 (m.); Borgmeier, 1959b: 348 (q.); Wheeler, G.C. 1949: 681 (l.).
- Combination in Oecodoma: Smith, F. 1858b: 184; Mayr, 1863: 438;
- combination in Atta: Roger, 1863b: 35.
- Subspecies of lebasii: Emery, 1890b: 55.
- Subspecies of cephalotes: Emery, 1913b: 259.
- Status as species: Smith, F. 1858b: 184; Roger, 1863b: 35; Mayr, 1863: 437; Mayr, 1865: 81; Forel, 1885a: 362; Dalla Torre, 1893: 152; Emery, 1894k: 58; Forel, 1899c: 33; Forel, 1908b: 40; Forel, 1908c: 348; Forel, 1912e: 179; Forel, 1913l: 239; Forel, 1914e: 10; Wheeler, W.M. 1923c: 5; Emery, 1924d: 353; Wheeler, W.M. 1925a: 36; Menozzi, 1927c: 268; Weber, 1938b: 205; Borgmeier, 1939: 422 (in list); Weber, 1941b: 127; Gonçalves, 1942: 346; Borgmeier, 1950d: 259; Weber, 1958a: 11; Borgmeier, 1959b: 345 (redescription); Kempf, 1972a: 26; Cherrett & Cherrett, 1989: 53; Bolton, 1995b: 75; Branstetter & Sáenz, 2012: 257; Fernández, et al. 2015: 137 (redescription); Guénard & Economo, 2015: 227; Fernández & Serna, 2019: 842.
- Senior synonym of erecta: Borgmeier, 1959b: 346; Kempf, 1972a: 27; Bolton, 1995b: 75; Fernández, et al. 2015: 137.
- Senior synonym of lebasii: Dalla Torre, 1893: 152; Forel, 1899c: 33, Emery, 1924d: 353; Kempf, 1972a: 27; Bolton, 1995b: 75; Fernández, et al. 2015: 137.
- Senior synonym of tonsipes: Borgmeier, 1959b: 346; Kempf, 1972a: 27; Bolton, 1995b: 75; Fernández, et al. 2015: 137.
- Distribution: Colombia, Costa Rica, Guatemala, Panama, Peru.
- erecta. Atta cephalotes var. erecta Santschi, 1929f: 92 (diagnosis in key) (w.) COSTA RICA.
- Type-material: holotype (?) worker.
- [Note: no indication of number of specimens is given.]
- Type-locality: Costa Rica: Santa Clara, Columbiana, 2.i.1924 (G.C. Wheeler).
- Type-depository: NHMB.
- Subspecies of cephalotes: Borgmeier, 1939: 422 (in list); Gonçalves, 1942: 345; Borgmeier, 1950d: 243.
- Junior synonym of colombica: Borgmeier, 1959b: 346; Kempf, 1972a: 27; Bolton, 1995b: 76; Fernández, et al. 2015: 137.
- lebasii. Atta lebasii Guérin-Méneville, 1844a: 422 (w.) COLOMBIA.
- Type-material: holotype (?) worker.
- [Note: no indication of number of specimens is given.]
- Type-locality: Colombia: (no further data) (Lebas).
- Type-depository: unknown (perhaps MNHN).
- [Note: according to Horn & Kahle, 1935: 99, Guérin’s Hymenoptera material was split between MNHN, MSNG, and ZSBS, but some are also present in NNML.]
- Emery, 1890b: 54 (q.m.).
- Combination in Oecodoma: Smith, F. 1858b: 184;
- combination in Atta: Roger, 1863b: 35.
- Status as species: Smith, F. 1858b: 184; Roger, 1863b: 35; Mayr, 1863: 438; Mayr, 1865: 81; Emery, 1890b: 54.
- Junior synonym of colombica: Dalla Torre, 1893: 152; Forel, 1899c: 33, Emery, 1924d: 353; Kempf, 1972a: 27; Bolton, 1995b: 76; Fernández, et al. 2015: 137.
- tonsipes. Atta columbica var. tonsipes Santschi, 1929f: 92 (diagnosis in key) (w.) PANAMA.
- Type-material: syntype workers (number not stated).
- Type-locality: Panama: Bellavista (W.M. Wheeler).
- Type-depository: NHMB (perhaps also MCZC).
- Subspecies of colombica: Borgmeier, 1939: 422 (in list); Weber, 1941b: 127; Gonçalves, 1942: 346; Borgmeier, 1950d: 243; Weber, 1958a: 12.
- Junior synonym of colombica: Borgmeier, 1959b: 346; Kempf, 1972a: 27; Bolton, 1995b: 77; Fernández, et al. 2015: 137.
Description
Karyotype
- See additional details at the Ant Chromosome Database.
- Explore: Show all Karyotype data or Search these data. See also a list of all data tables or learn how data is managed.
- n = 11, 2n = 22, karyotype = 12M+6SM+4A (Panama) (Murakami et al., 1998; Barros et al., 2014) (Atta columbica is a writing mistake).
References
- Guérin-Méneville, F. E. 1844a. Iconographie du règne animal de G. Cuvier, ou représentation d'après nature de l'une des espèces les plus remarquables, et souvent non encore figurées, de chaque genre d'animaux. Insectes. Paris: J. B. Baillière, 576 pp.
- Armitage, S., Boomsma, J., Bear, B. 2010. Diploid male production in a leaf-cutting ant. Ecological Entomology 35, 175–182 (doi:10.1111/j.1365-2311.2009.01167.x).
- Baer, B. 2011. The copulation biology of ants (Hymenoptera: Formicidae). Myrmecological News 14: 55-68.
- Barrera, C.A., Sosa-Calvo, J., Schultz, T.R., Rabeling, C., Bacci, M., Jr 2021. Phylogenomic reconstruction reveals new insights into the evolution and biogeography of Atta leaf-cutting ants (Hymenoptera: Formicidae). Systematic Entomology 47: 13-35 (doi:10.1111/syen.12513).
- Barros, L.A.C., Teixeira, G.A., de Aguiar, J.A.C., Mariano, C.D.S.F., Delabie, J.H.C., Pompolo, S.D.G. 2014. Banding Patterns of Three Leafcutter Ant Species of the Genus Atta (Formicidae: Myrmicinae) and Chromosomal Inferences. Florida Entomologist 97(4): 1694-1701.
- Bochynek, T., J. L. Tanner, B. Meyer, and M. Burd. 2017. Parallel foraging cycles for different resources in leaf-cutting ants: a clue to the mechanisms of rhythmic activity. Ecological Entomology. 42:849-852. doi:10.1111/een.12437
- Bustamante, S., Amarillo-Suárez, A. 2019. Foraging plasticity of Atta cephalotes (Hymenoptera: Formicidae) in response to temperature differences between forest and pasture. Revista de Biología Tropical 67(4), 963-974.
- Cardoso, D. C., Cristiano, M. P. 2021. Karyotype diversity, mode, and tempo of the chromosomal evolution of Attina (Formicidae: Myrmicinae: Attini): Is there an upper limit to chromosome number? Insects 1212, 1084 (doi:10.3390/insects12121084).
- Dahan, R.A., Grove, N.K., Bollazzi, M., Gerstner, B.P., Rabeling, C. 2021. Decoupled evolution of mating biology and social structure in Acromyrmex leaf-cutting ants. Behavioral Ecology and Sociobiology 76, 7 (doi:10.1007/s00265-021-03113-1).
- de Bekker, C., Will, I., Das, B., Adams, R.M.M. 2018. The ants (Hymenoptera: Formicidae) and their parasites: effects of parasitic manipulations and host responses on ant behavioral ecology. Myrmecological News 28: 1-24 (doi:10.25849/myrmecol.news_028:001).
- Dijkstra, M. B. and J. J. Boomsma. 2006. Are workers of Atta leafcutter ants capable of reproduction? Insectes Sociaux. 53(2):136-140. doi:10.1007/s00040-005-0848-3
- Dijkstra, M. B., D. R. Nash, and J. J. Boomsma. 2005. Self-restraint and sterility in workers of Acromyrmex and Atta leafcutter ants. Insectes Sociaux. 52(1):67-76. doi:10.1007/s00040-004-0775-8
- Farias, A.P., Camargo, R.da S., Caldato, N., Forti, L.C. 2020. Nest architecture development of grass-cutting ants, Atta capiguara (Hymenoptera: Formicidae). Revista Brasileira de Entomologia 64, e20200034 (doi:10.1590/1806-9665-rbent-2020-0034).
- Fleischmann, P.N., Grob, R., Rössler, W. 2020. Magnetoreception in Hymenoptera: importance for navigation. Animal Cognition (doi:10.1007/s10071-020-01431-x).
- Horna-Lowell, E., Neumann, K.M., O’Fallon, S., Rubio, A., Pinter-Wollman, N. 2021. Personality of ant colonies (Hymenoptera: Formicidae) – underlying mechanisms and ecological consequences. Myrmecological News 31: 47-59 (doi:10.25849/MYRMECOL.NEWS_031:047).
- Hull-Sanders, H. M., and Howard, J. J. 2003. Impact of Atta colombica colonies on understory vegetation and light availability in a Neotropical forest. Biotropica 35(3): 441–445
- Jacobs, S. 2020. Population genetic and behavioral aspects of male mating monopolies in Cardiocondyla venustula (Ph.D. thesis).
- Lau, M.K., Ellison, A.M., Nguyen, A., Penick, C., DeMarco, B., Gotelli, N.J., Sanders, N.J., Dunn, R.R., Helms Cahan, S. 2019. Draft Aphaenogaster genomes expand our view of ant genome size variation across climate gradients. PeerJ 7, e6447 (doi:10.7717/PEERJ.6447).
- Li, H., Young, S.E., Poulsen, M., Currie, C.R. 2020. Symbiont-mediated digestion of plant biomass in fungus-farming insects. Annual Review of Entomology 66:16.1–16.20 (doi:10.1146/annurev-ento-040920-061140).
- Matte, A., Billen, J. 2012. Flight muscle histolysis in Lasius niger queens. Asian myrmecology 13, e013003 (doi:10.20362/am.013003).
- Mueller, U.G., Ishak, H.D., Bruschi, S.M., Smith, C.C., Herman, J.J., Solomon, S.E., Mikheyev, A.S., Rabeling, C., Scott, J.J., Cooper, M., Rodrigues, A., Ortiz, A., Brandão, C.R.F., Lattke, J.E., Pagnocca, F.C., Rehner, S.A., Schultz, T.R., Vasconcelos, H.L., Adams, R.M.M., Bollazzi, M., Clark, R.M., Himler, A.G., LaPolla, J.S., Leal, I.R., Johnson, R.A., Roces, F., Sosa-Calvo, J., Wirth, R., Bacci, M. 2017. Biogeography of mutualistic fungi cultivated by leafcutter ants. Molecular Ecology 26, 6921–6937 (doi:10.1111/mec.14431).
- Mueller, U.G., Schultz, T.R., Currie, C.R., Adams, R.M.M., Malloch, D. 2001. The origin of the attine ant-fungus mutualism. The Quarterly Review of Biology 76, 169-197.
- Narendra, A., Ramirez-Esquivel, F., Ribi, W.A. 2016. Compound eye and ocellar structure for walking and flying modes of locomotion in the Australian ant, Camponotus consobrinus. Scientific Reports 6, 22331 (doi:10.1038/srep22331).
- Nehring, V., Boomsma, J.J., d'Ettorre, P. 2012. Wingless virgin queens assume helper roles in Acromyrmex leaf-cutting ants. Current Biology 22, R671–R673 (doi:10.1016/j.cub.2012.06.038).
- Pokorny, T., Sieber, L.-M., Hofferberth, J.E., Bernadou, A., Ruther, J. 2020. Age-dependent release of and response to alarm pheromone in a ponerine ant. The Journal of Experimental Biology 223, jeb218040. (doi:10.1242/JEB.218040).
- Ramalho, M.de O., Martins, C., Morini, M.S.C., Bueno, O.C. 2020. What can the bacterial community of Atta sexdens (Linnaeus, 1758) tell us about the habitats in which this ant species evolves? Insects 11, 332. (doi:10.3390/INSECTS11060332).
- Ronque, M.U.V., Lyra, M.L., Migliorini, G.H., Bacci, M., Oliveira, P.S. 2020. Symbiotic bacterial communities in rainforest fungus-farming ants: evidence for species and colony specificity. Scientific Reports 10, 10172 (doi:10.1038/S41598-020-66772-6).
- Toledo-Hernández, E., Hernández-Flores, J., Sotelo-Leyva, C., Alvear-García, A., Peña-Chora, G. 2021. A new enemy of Apis mellifera (Hymenoptera: Apidae): First report of Nomamyrmex esenbeckii (Hymenoptera: Formicidae) attacking honey bee colonies. Journal of Apicultural Research, 1–2 (doi:10.1080/00218839.2021.1987740).
References based on Global Ant Biodiversity Informatics
- Anderson, C. and J.L.V. Jadin. 2001. The adaptive benefit of leaf transfer in Atta colombica. Insectes Sociaux 48:404-405
- Baer, B., S. P. A. den Boer, D. J. C. Kronauer, D. R. Nash and J. J. Boomsma. 2009. Fungus gardens of the leafcutter ant Atta colombica function as egg nurseries for the snake Leptodeira annulata. Insectes Sociaux 56(3):289-291
- Borgmeier T. 1939. Nova contribuição para o conhecimento das formigas neotropicas (Hym. Formicidae). Revista de Entomologia (Rio de Janeiro) 10: 403-428.
- Borgmeier T. 1950. Atta-Studien (Hym. Formicidae). Memórias do Instituto Oswaldo Cruz. Rio de Janeiro 48: 265-292.
- Borgmeier T. 1950. Estudos sôbre Atta (Hym. Formicidae). Memórias do Instituto Oswaldo Cruz. Rio de Janeiro 48: 239-263.
- Bustos H., J. 1994. Contribucion al conocimiento de al fauna de hormigas (Hymenoptera: Formicidae) del occidente del Departamento de Narino (Colombia). Bol. Mus. Ent. Univ. Valle 2(1,2):19-30
- Davidson D. W., S. C. Cook, R. R. Snelling and T. H. Chua. 2003. Explaining the Abundance of Ants in Lowland Tropical Rainforest Canopies. Science 300: 969-972.
- Davidson, D.W. 2005. Ecological stoichiometry of ants in a New World rain forest. Oecologia 142:221-231
- Emery C. 1890. Studii sulle formiche della fauna neotropica. Bull. Soc. Entomol. Ital. 22: 38-8
- Emery C. 1894. Estudios sobre las hormigas de Costa Rica. Anales del Museo Nacional de Costa Rica 1888-1889: 45-64.
- Fernández F., E. E. Palacio, W. P. Mackay, and E. S. MacKay. 1996. Introducción al estudio de las hormigas (Hymenoptera: Formicidae) de Colombia. Pp. 349-412 in: Andrade M. G., G. Amat García, and F. Fernández. (eds.) 1996. Insectos de Colombia. Estudios escogidos. Bogotá: Academia Colombiana de Ciencias Exactas, Físicas y Naturales, 541 pp
- Fernández, F. and S. Sendoya. 2004. Lista de las hormigas neotropicales. Biota Colombiana Volume 5, Number 1.
- Forel A. 1908. Fourmis de Costa-Rica récoltées par M. Paul Biolley. Bulletin de la Société Vaudoise des Sciences Naturelles 44: 35-72.
- Forel A. 1912. Formicides néotropiques. Part II. 3me sous-famille Myrmicinae Lep. (Attini, Dacetii, Cryptocerini). Mémoires de la Société Entomologique de Belgique. 19: 179-209.
- Gonçalves C. R. 1942. Contribuiça~o para o conhecimento do gênero Atta Fabr., das formigas saúvas. Bol. Soc. Bras. Agron. 5: 333-358.
- Helmkampf, M., J. Gadau and H. Feldhaar. 2008. Population- and sociogenetic structure of the leaf-cutter ant Atta colombica (Formicidae, Myrmicinae). Insectes Sociaux 55(4):434-442.
- INBio Collection (via Gbif)
- Johnson R. Personnal Database. Accessed on February 5th 2014 at http://www.asu.edu/clas/sirgtools/resources.htm
- Kaspari, M. and M.D. Weiser. Ant Activity along Moisture Gradients in a Neotropical Forest Ant Activity along Moisture Gradients in a Neotropical Forest. Biotropica, Vol. 32, No. 4a (Dec., 2000), pp. 703-711
- Kempf, W.W. 1972. Catalago abreviado das formigas da regiao Neotropical (Hym. Formicidae) Studia Entomologica 15(1-4).
- Kooij P. W., B. M. Dentinger, D. A. Donoso, J. Z. Shik, and E. Gaya. 2018. Cryptic diversity in Colombian edible leaf-cutting ants (Hymenoptera: Formicidae). Insects 9: 191.
- Kost, C., E. Gama de Oliveira, T.A. Knoch, R. Wirth. 2005. Spatio-Temporal Permanence and Plasticity of Foraging Trails in Young and Mature LeafCutting Ant Colonies (Atta spp.). Journal of Tropical Ecology 21(6):677-688
- Kusnezov N. 1953. La fauna mirmecológica de Bolivia. Folia Universitaria. Cochabamba 6: 211-229.
- Longino J. et al. ADMAC project. Accessed on March 24th 2017 at https://sites.google.com/site/admacsite/
- Menozzi C. 1927. Formiche raccolte dal Sig. H. Schmidt nei dintorni di San José di Costa Rica. Entomologische Mitteilungen. Berlin-Dahlem. 16: 266-277.
- Murakami T., A. Fujiwara,and M. C. Yoshida. 1999. Cytogenetics of ten ant species of the tribe Attini (Hymenoptera, Formicidae) in Barro Colorado Island, Panama. Chromosome Science 2(3): 135-139.
- Powell, S. and E. Clark. 2004. Combat between large derived societies: A subterranean army ant established as a predator of mature leaf-cutting ant colonies. Insectes Sociaux 51(4):342-351.
- Sturup, M., S.P.A. den Boer, D.R. Nash, J.J. Boomsma and B. Baer. 2011. Variation in male body size and reproductive allocation in the leafcutter ant Atta colombica: estimating variance components and possible trade-offs. Insectes Sociaux 58:47-55
- Ulloa Chacon P., M. L. Baena, J. Bustos, R. C. Aldana, J. A. Aldana, and M. A. Gamboa. 1996. Fauna de hormigas del departamento del Valle del Cauca (Colombia). Pp. 413-451. In Andrade-C M. G., G. Amat Garcia, and F. Fernandez. Insectoss de Colombia, estudios escogidos.
- Weber N. A. 1941. The biology of the fungus-growing ants. Part VII. The Barro Colorado Island, Canal Zone, species. Rev. Entomol. (Rio J.) 12: 93-130.
- Weber N. A. 1958. Nomenclatural notes on Proatta and Atta (Hym.: Formicidae). Entomological News 69: 7-13.
- Weber N. A. 1968. The Panamanian Atta species (Hymenoptera: Formicidae). Proceedings of the Entomological Society of Washington 70: 348-350.
- Weber N. A. 1969. Ecological relations of three Atta species in Panama. Ecology 50: 141-147.
- Pages using DynamicPageList3 parser function
- Photo Gallery
- Need species key
- Tropical
- Phorid fly Associate
- Host of Apocephalus colombicus
- Host of Apocephalus attophilus
- Host of Eibesfeldtphora pala
- Fungus Associate
- Host of Ophiocordyceps subramanianii
- Karyotype
- Species
- Extant species
- Formicidae
- Myrmicinae
- Attini
- Atta
- Atta colombica
- Myrmicinae species
- Attini species
- Atta species