Wasmannia auropunctata
| Wasmannia auropunctata | |
|---|---|

| |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Arthropoda |
| Class: | Insecta |
| Order: | Hymenoptera |
| Family: | Formicidae |
| Subfamily: | Myrmicinae |
| Tribe: | Attini |
| Genus: | Wasmannia |
| Species: | W. auropunctata |
| Binomial name | |
| Wasmannia auropunctata (Roger, 1863)
| |
| Synonyms | |
| |


| Common Name | |
|---|---|
| Little Fire Ant | |
| Language: | English |
An infamous tramp ant that has been introduced to many areas outside of its native range, and is one of the 100 worst invasive species in the world (IUCN/SSC Invasive Species Specialist Group). It has a notoriously strong sting for an ant of its size. Longino & Fernández (2007) offered the following account of one myrmecologist's experience with this ants famous stinging abilities: These are extremely tiny ants, barely visible in the field. When the senior author first began studying ants in Costa Rica, he was at first puzzled about Wasmannia. By literature accounts Wasmannia was reputed to have a terrible sting, but he had been collecting them for months in Corcovado National Park without ever experiencing the famous sting. One day he was collecting from a populous nest and some workers made it up to the soft skin of his inner forearm and began to sting. The sting was definitely noticeable, about as severe as a fire ant (i. e., Solenopsis geminata) but inordinately strong for an ant that could barely be seen! Workers are so small they cannot sting through the thicker skin of the hands.
| At a Glance | • Highly invasive • Parthenogenetic • Supercolonies |
Identification
Workers of Wasmannia auropunctata have a strongly quadrate petiolar node. The anterior face of the node is sharply differentiated from both the peduncle and the dorsal face of the node, meeting both at nearly right angles, and forming a strongly step-like profile. This is a highly distinctive feature that easily distinguishes auropunctata workers from all other Wasmannia species. (Longino & Fernández 2007)
There is considerable variation in the development of head sculpture and color.
Wetterer and Hita-Garcia (2015) - Tetramorium caldarium workers are similar in body size, proportions, and color to another tramp ant species, Wasmannia auropunctata. Wasmannia auropunctata, however, is slightly smaller, has longer setae on the face and dorsum, longer propodeal spines, and a more rectangular petiole in side view. Another important key difference is the presence of a two-segmented antennal club in W. auropunctata compared to the three segmented club of T. caldarium. The latter also has very weak frontal carinae and reduced antennal scrobes while both of these characters are strongly developed in W. auropunctata.

{kind=link}
Keys including this Species
Distribution
Latitudinal Distribution Pattern
Latitudinal Range: 27.158° to -64.36°.
| North Temperate |
North Subtropical |
Tropical | South Subtropical |
South Temperate |
- Source: AntMaps
Distribution based on Regional Taxon Lists
Afrotropical Region: Cameroun, Gabon, Sierra Leone.
Australasian Region: Australia, New Caledonia.
Indo-Australian Region: Hawaii, Philippines, Solomon Islands, Vanuatu.
Nearctic Region: Canada, United States.
Neotropical Region: Antigua and Barbuda, Argentina, Aruba, Barbados, Belize, Bermuda, Bolivia, Brazil, Colombia, Costa Rica, Cuba (type locality), Dominican Republic, Ecuador, French Guiana, Galapagos Islands, Grenada, Guadeloupe, Guatemala, Haiti, Honduras, Lesser Antilles, Mexico, Netherlands Antilles, Panama, Paraguay, Peru, Puerto Rico, Saint Lucia, Uruguay, Venezuela.
Palaearctic Region: Cyprus, Israel, Italy, Spain.
Distribution based on AntMaps
Distribution based on AntWeb specimens
Check data from AntWeb
Countries Occupied
| Number of countries occupied by this species based on AntWiki Regional Taxon Lists. In general, fewer countries occupied indicates a narrower range, while more countries indicates a more widespread species. |
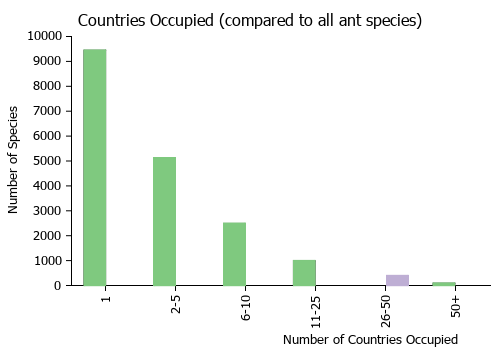
|
Biology
Longino & Fernández (2007) - Wasmannia auropunctata is a widespread pest ant (Clark et al., 1982; De Souza et al., 1998; Fabres & Brown, 1978; Jourdan, 1997; Lubin, 1984; Ulloa Chacon & Cherix, 1990; Williams, 1994; Wetterer & Porter, 2003). In its presumed native range it occurs from Argentina to Mexico (Kempf, 1972; Wetterer & Porter, 2003). Its introduced range includes the Galapagos Islands, West Africa (Gabon, Cameroon, and possibly the Republic of Congo and the Democratic RepubIic of Congo), Melanesia (New Caledonia, Solomon Islands, Vanuatu, and possibly Tuvalu), Polynesia (Wallis and Futuna and Hawaii), parts of the US (Florida and possibly California), and subtropical Atlantic islands (the Bahamas and Bermuda) (Wetterer & Porter, 2003). It is widespread on Caribbean islands, but it is unclear whether these are long-term native populations or recent introductions (Wetterer & Porter, 2003).
The species is remarkably catholic in its habitat preference. It is common in habitats ranging from wet to dry and from early successional to mature. In an elevational gradient of mature wet forest on the Atlantic slope of Costa Rica (the Barva Transect, from La Selva Biological Station to 2000 m elevation on the slope of Volcan Barva) it is abundant at 50 m and 500 m elevations, but nearly absent at 1070 m . In the lowland habitats where it is abundant, it occurs in leaf litter on the forest floor and at all levels in the vegetation.
Although it occurs frequently in samples from mature forest habitats in Costa Rica, it is never so abundant in those habitats that it is noticeable as a pest or appears to be displacing other native species (Tennant, 1994; McGlynn & Kirksey, 2000; pers. obs.). In contrast, in certain agricultural habitats (banana plantations) and in parts of the tropics where it has been introduced it becomes super-abundant, with negative impacts on native species and human comfort (Clark et al., 1982; Wetterer & Porter, 2003). In dry-forest fragments in Colombia there is a negative correlation between W. auropunctata abundance and overall ant diversity (Armbrecht & Ulloa Chacon, 2003). Where introduced in New Caledonia it invades dense native forest and displaces native ants (Le Breton et al., 2003). Behavioral tests and cuticular hydrocarbon analysis show that W. auropunctata is multicolonial in its native range in Brazil, unicolonial where introduced in New Caledonia (Errard et al., 2002).
Nests can be almost anywhere: in rolled leaves or dead sticks in the leaf litter, under stones, in rotten wood, in hollow stems suspended above the ground, in ant-plant domatia, and under epiphytes. Workers are omnivorous scavengers and predators and can rapidly recruit to food. Colonies are polygynous and it is never clear where colony boundaries are. Dozens of dealate queens may be found together in nests. Males are rare but do occasionally occur.
Surprisingly, the chemical and toxicological nature of the venom of W. auropunctata has not been investigated. Howard et al. (1982) discovered an alkylpyrazine compound in the mandibular glands, which acted as an attractant to conspecifics and a repellent to heterospecifics. They speculated that the workers might apply the mandibular gland product as an irritating secretion, augmenting the defensive properties of the venomous sting. It would be interesting to investigate whether the venom alone is the powerful agent in this small ant, or if the strong burning sensation is a synergetic effect of venom plus mandibular gland product.
Cuezzo et al. (2015) - Kusnezov (1952) hypothesized parthenogenetic reproduction by this species given the high prevalence of colonies containing only females, this was later confirmed by Fournier (et al., 2005). The LFA displays extraordinary reproductive polymorphism with both regular sexual and unusual clonal populations (Foucaud et al., 2007). This rare type of clonality was also recently reported for the invasive longhorn crazy ant Paratrechina longicornis (Pearcy et al., 2011). This unusual reproductive system is postulated to be responsible for unicoloniality, in which individuals from different nests form a large supercolony, an attribute that probably contributes to the success as an invader (Orivel et al., 2009).
Although numerous studies were conducted on the LFA in northern South America (Ulloa-Chacón & Cherix, 1990, de Souza et al., 1998, Foucaud et al., 2009, Orivel et al., 2009), little is known about populations at the southern limit of its native range (Kuznezov, 1952, Rey et al. 2012, Calcaterra et al., 2012).
Bertelsmeier et al. (2015a, b) examined elements of interspecific aggression, and food resource discovery and dominance, between this species and several other highly invasive ants. In laboratory assays Wasmannia auropunctata was the most tenacious in direct interactions. Of the group of four species that were found to be aggressive, W. auropunctata was found to be the slowest discoverer of and recruited the fewest workers to food in a laboratory arena experiment.
Foraging
Rosumek (2017) examined trophic and temporal niches of ants in a secondary Atlantic forest (Florianópolis, south Brazil; 27°31′38″S 48°30′15″W / 27.52722°S 48.50417°W), between December 2015 and January 2016. For this species: A large body of knowledge describes how W. auropunctata dominate habitats and displace other ants, which often happens when the species is introduced or, within its native range, in crops and other open/disturbed areas. The species is portrayed feeding virtually on everything: scavenging; preying on small and large arthropods; collecting diversified plant parts; visiting extra-floral nectaries and tending honeydew-producing insects (Creighton, 1950; Kusnezov, 1952; Smith, 1954; Smith, 1965; Fabres & Brown, 1978; Clark et al., 1982; Deyrup et al., 2000; Wetterer & Porter, 2003; Longino & Fernández, 2007). Some of these authors suggest that honeydew is their main resource, such as Clark et al. (1982).
A comparatively small amount of information suggests that, inside forests within its native range, the species is not nearly as dominant (Majer & Delabie, 1999; Longino & Fernández, 2007). Very little is known about W. auropunctata habits in this context. Using generic baits, Orivel et al. (2009) showed a steep decline in bait use and nest density within a gradient from open areas to undisturbed forest. In Atlantic forest, ca. 1400 km north of the present study site, Santana et al. (2013) qualitatively showed it interacting with seven non- myrmecophorous diaspores on the ground.
In light of this previous work, it was really surprising to find the species to be a strict specialist in feces. In fact, it was the only species in this study that used a single resource. It was a comparatively frequent species, but appeared always in low numbers and was not collected in pitfalls. This result differs from the widespread use of fleshy diaspores found in Santana et al. (2013), and also from the use of baits in Orivel et al. (2009). The latter authors suggested that abiotic factors play a role in the ecological shift of W. auropunctata from open to forest areas. A physiological constraint related to environmental conditions (e.g. temperature) could explain why the species has a limited role inside forests, and why, in a higher latitude, it shifts to a resource less preferred by other species.
This species showed no preference in its time of foraging.
Regional Information
Argentina
Cuezzo et al. (2015) - W. auropunctata is widely distributed in Argentina; however, it is common only in anthropic habitats, mostly in northeastern Argentina (Rey et al., 2012; L.A.C, unpublished data). Nests are usually found in urban areas under stones, around or inside tree trunks, and under sidewalks. W. auropunctata was found in Lozano (34º51´S, 59º03´W, 45m, Francisco Sola coll.) in Buenos Aires province, representing the southernmost record reported so far for this species. Unexpectedly, it was also found for the first time in the arid desert of the Monte ecoregion from Salta province to the locality of Anillaco (28°49’S, 66°56’W, Adriana Aranda coll.) in La Rioja province, and up to 2125 m elevation in the locality of Tumbaya (23º51’S, 65º28’W), Puna ecoregion, Jujuy province. Wasmannia auropunctata is uncommon in minimally disturbed native habitats where ant assemblages are typically more diverse compared with anthropic habitats (L.A.C., unpublished data). Though scarce in native forests in northern Argentina (L.A.C., unpublished data). W. auropunctata was only common in one type of agricultural habitat (banana plantations) in the Jujuy (Yungas) and Formosa (Chaco) provinces.
Two specimens of gynandromorphs were found in a nest of W. auropunctata with putatively sexual castes reproducing clonally, in Colón (32º14´S, 58º08´W), Entre Ríos province, Argentina. This is the first record of gynandromorphs occurring naturally in a LFA colony, however, additional gynandromorphs have been found recently in natural populations of W. auropunctata in São Sebastian, Brazil (LAC, unpublished data) and also found by F. Cuezzo in series of W. auropunctata collected by N. Kusnezov and deposited in IFML. Gynandromorphs previously have been found in clonal LFA colonies that were exposed to temperatures of more than 40°C in the laboratory (Olivier Rey, personal communication). They could have been induced in the laboratory during embryonic development by the effect of the extremely high rearing temperature. Gynandromorphism is common in Hymenoptera (ants: Jones & Phillips, 1985; bees: Wcislo et al., 2004). These sexual mosaics have been reported in more than 40 ant species (Jones &Phillips, 1985) but, to date, their production has only been explained in two cases: sub-lethally high breeding temperature in the pharaoh’s ant, Monomorium pharaonis (L.) (Berndt & Kremer, 1982), and Wolbachia infections in the isopod Armadillidium vulgare (Rigaud & Juchault, 1993).
The wide distribution and relatively high abundance of W. auropunctata could be explained by the fact it has the largest and most fecund queen within the genus (Kusnezov, 1952; Longino & Fernández, 2007). However, the higher abundance of W. auropunctata in disturbed habitats, mostly in central-eastern Argentina, seems to be more related with the fact that sexual castes are produced almost exclusively by clonal reproduction (clonal populations) in this region (Rey et al. 2012, LAC, unpublished data).
Clonal populations of W. auropunctata were found up to 2125 m elevation in the locality of Tumbaya, Jujuy. This was surprising because it had been previously found only up to approximately 1070 m elevation (Wetterer & Porter, 2003). The discovery of W. auropunctata in the Puna ecosystem (Tumbaya, Jujuy) under extremely cold and dry conditions (annual rainfall 179 mm and mean and minimum temperatures of 8.1ºC and -8ºC in July, the coldest month, De Fina, 1992) was unexpected. According to Kusnezov (1952), W. auropunctata was not able to resist prolonged drought. However, our finding supports laboratory evidence that clonal populations present in Argentina are strongly adapted to very low temperatures and probably also to low levels of humidity in environments such as the Puna. This record represents both the highest altitude and the most severe natural environmental conditions reported so far for this species.
Chifflet et al (2018) - Evidence suggests a recent southern range expansion during the last 60 years reaching central Argentina. This supercolonial ant species has a polymorphic reproductive system. Some populations, mostly found in undisturbed natural environments, are characterised by a classical sexual haplodiploid reproductive system. In other populations, which mainly occur in human-modified habitats, diploid queens and haploid males are produced clonally while workers are produced sexually. Here we studied the association between the recent southern range expansion of W. auropunctata in relation to human activity and clonality. We carried out an extensive survey within the southern limit of the species' native distribution and characterised the type of habitat where populations were found. Moreover, we genetically determined the type of reproductive system in 35 populations by genotyping at 12 microsatellite loci a total of 191 reproductive individuals (i.e. queens and/or males). Clonality was the most common reproductive system, occurring in 31 out of 35 populations analysed. All the populations found in the recently colonised area in central Argentina were clonal and established in human-modified habitats, suggesting that clonality together with human activity might have facilitated the southwards expansion of W. auropunctata.
Cameroon
Mbenoun Masse et al. (2017) - W. auropunctata is present in Cameroon where it invades mainly disturbed habitats such as urban and rural areas. Despite being present for more than four decades, its impact on local ant diversity was unknown. We studied the impact of W. auropunctata in three disturbed habitats located in rural and urban areas. We monitored ant diversity in both invaded and noninvaded zones using baits, pitfall traps and visual catch in quadrats. We collected 28 species in urban area and 64 in rural area. We found a negative impact of the little fire ant on abundance, richness and community composition of the local ant species. In invaded zone, W. auropunctata made up 97.72% and 99.96% of all ant fauna and ant species richness decreased to 7 and 2 in urban and rural area, respectively.
Florida
Deyrup, Davis & Cover (2000): A common species in south Florida, occurring sporadically as far north as Marion County. Nests are in leaf litter, in rotten wood or hollow twigs on the ground, at the bases of trees, and frequently in flower pots. Habitats that are wet or dry, shaded or open, are ail acceptable. On some sites the populations are huge, the massed workers forming golden patches on the underside of any object that has been left on the ground, but it is generally a minor species in the fauna. Where it is most abundant, it is easy to see how this ant could become a major pest (because of its powerful sting) and reduce populations of native species, as it has in the Galapagos (Clark et al. 1982, Lubin 1984). We have recently found sites on the island of New Providence, Bahamas, with huge concentrations of W. auropunctata. The factors that trigger or support these outbreaks are unknown. At two outbreak sites in south Florida there are unusual concentrations of plants that produce large amounts of extra-floral nectar, and are visited day and night by hordes of W. auropunctata. It may be that this resource has tipped the balance in favor of W. auropunctata and caused these very local outbreaks.
This species is a general scavenger, and seems quite dependent on extra-floral nectaries. Pest status: this species packs a sting out of all proportion to its size, and humans are frequently stung by ants that have fallen out of trees, or are foraging across lawn chairs, or floating on the surface of swimming pools. Foraging columns often move for many yards, and may invade homes, where stray ants sting the occupants. First published Florida record: Smith 1929; earlier specimens: 1924.
Association with Other Organisms
 Explore: Show all Associate data or Search these data. See also a list of all data tables or learn how data is managed.
Explore: Show all Associate data or Search these data. See also a list of all data tables or learn how data is managed.
- This species is a host for the eucharitid wasp Orasema minutissima (a parasite) (Mann, 1918; Heraty, 1994; Wetterer & Porter, 2003; Burks et al., 2018; Baker et al., 2019; Heraty et al., 2021; Universal Chalcidoidea Database) (primary host).
Flight Period
| X | X | X | X | X | X | X | X | ||||
| Jan | Feb | Mar | Apr | May | Jun | Jul | Aug | Sep | Oct | Nov | Dec |
Source: Kaspari et al., 2001.
- Check details at Worldwide Ant Nuptial Flights Data, AntNupTracker and AntKeeping.
- Explore: Show all Flight Month data or Search these data. See also a list of all data tables or learn how data is managed.
Life History Traits
- Queen number: polygynous (Holldobler & Wilson, 1977; Frumhoff & Ward, 1992)
Castes
Longino & Fernández (2007) - There appear to be two size classes of queens. Queens with smaller heads include ten from various sites in Costa Rica, including La Selva Biological Station and the Penas Blancas Valley, one from Jamaica, and one from Venezuela. Queens with large heads are all from the Atlantic slope of Costa Rica. Three are from La Selva (two from different Winkler samples of sifted leaf litter from the forest floor, one from a small nest under an epiphyte mat in an old treefall) and one is an alate queen found in a Cecropia sapling near Volcan Arenal. Thus the small-headed and big-headed forms are broadly sympatric in Costa Rica. Among the small-headed queens four are definitively associated with workers from the same colony, and among the big-headed queens one is associated with workers. Others have workers doubtfully associated (together in the same Winkler sample). We can discern no differences in workers associated with the two types of queens. The cause of the two size classes of queens is unknown, but could reflect either differences between cryptic species or intraspecific polymorphism.
An aberrant worker form is frequently encountered in large samples of W. auropunctata. These aberrant workers have the head grossly swollen. The entire head is more spherical than normal, as if the head were inflated like a balloon. The rest of the body is little different from a normal worker. These aberrant workers are occasionally encountered in Winkler samples that contain hundreds or thousands of auropunctata workers.
Cuezzo and Calcaterra (2015) - Two sympatric sizes of gynes were described in Costa Rica on the basis of their head size (Longino & Fernández, 2007) and could represent either differences between cryptic species or intraspecific polymorphisms. Gyne variation in head size was not observed in Argentina. Argentinean queens were most similar to the small headed form found in Costa Rica, Jamaica, and Venezuela (Longino & Fernández, 2007). According to Kusnezov (1952), the worker size of populations of W. auropunctata from Misiones province differ from the size of workers found in other populations elsewhere in Argentina; however, we have not observed such variation.
Worker
Images from AntWeb
   
| |
| Worker. Specimen code casent0005665. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by UCDC, Davis, CA, USA. |
   
| |
| Worker. Specimen code casent0039797. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by CAS, San Francisco, CA, USA. |
   
| |
| Worker. Specimen code casent0106114. Photographer Michael Branstetter, uploaded by California Academy of Sciences. | Owned by UCDC, Davis, CA, USA. |
   
| |
| Worker. Specimen code casent0173249. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by CDRS, Galapagos, Ecuador. |
   
| |
| Worker. Specimen code casent0178667. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by MIZA, Maracay, Venezuela. |
   
| |
| Worker. Specimen code casent0179473. Photographer Erin Prado, uploaded by California Academy of Sciences. | Owned by CAS, San Francisco, CA, USA. |
 
| |
| Worker. Specimen code casent0178173. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by ALWC, Alex L. Wild Collection. |
Queen
Images from AntWeb
    
| |
| Queen (alate/dealate). Specimen code casent0102747. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by ABS, Lake Placid, FL, USA. |
Male
Images from AntWeb
    
| |
| Male (alate). Specimen code casent0102748. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by ABS, Lake Placid, FL, USA. |
     
| |
| Male (alate). Specimen code casent0173250. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by CDRS, Galapagos, Ecuador. |
Nomenclature
The following information is derived from Barry Bolton's Online Catalogue of the Ants of the World.
- auropunctata. Tetramorium auropunctatum Roger, 1863a: 182 (w.q.m.) CUBA. Wheeler, G.C. & Wheeler, J. 1954d: 444 (l.). Combination in Ochetomyrmex: Forel, 1886b: xlix; in Wasmannia: Forel, 1893g: 383. Senior synonym of atomum: Wheeler, W.M. 1922a: 912; of glabra: Kempf, 1964e: 66; of panamana: Brown, 1948d: 102; of australis, laevifrons, nigricans, obscura, pulla, rugosa: Longino & Fernández, 2007: 276.
- rugosa. Ochetomyrmex auropunctatus var. rugosus Forel, 1886b: xlix (w.) GUATEMALA. Santschi, 1929d: 299 (q.m.). Combination in Wasmannia: Forel, 1901c: 128. Subspecies of auropunctata: Emery, 1888c: 353. Junior synonym of auropunctata: Longino & Fernández, 2007: 276.
- australis. Wasmannia auropunctata var. australis Emery, 1894c: 193 (w.) BRAZIL. Junior synonym of auropunctata: Longino & Fernández, 2007: 276.
- laevifrons. Wasmannia auropunctata var. laevifrons Emery, 1894c: 193 (w.) BOLIVIA. Wheeler, W.M. 1925a: 36 (q.). Junior synonym of auropunctata: Longino & Fernández, 2007: 276.
- nigricans. Wasmannia auropunctata var. nigricans Emery, 1906c: 160 (w.) PARAGUAY. Junior synonym of auropunctata: Longino & Fernández, 2007: 276.
- obscura. Wasmannia auropunctata var. obscura Forel, 1912g: 1 (w.) COLOMBIA. Junior synonym of auropunctata: Longino & Fernández, 2007: 276.
- atomum. Xiphomyrmex atomum Santschi, 1914d: 370 (w.) GABON. Combination in Wasmannia: Santschi, 1916b: 504. Subspecies of auropunctata: Santschi, 1916b: 504. Junior synonym of auropunctata: Wheeler, W.M. 1922a: 912.
- weiseri. Wasmannia sulcaticeps var. weiseri Forel, 1914d: 281 (q.m.) ARGENTINA. Junior synonym of auropunctata: Longino & Fernández, 2007: 281.
- glabra. Wasmannia glabra Santschi, 1931c: 272 (q.) FRENCH GUIANA. Junior synonym of auropunctata: Kempf, 1964e: 66.
- pulla. Wasmannia auropunctata st. pulla Santschi, 1931c: 272 (w.) PANAMA. Junior synonym of auropunctata: Longino & Fernández, 2007: 277.
- panamana. Hercynia panamana Enzmann, J. 1947a: 44, pl. 5 (w.q.) PANAMA. Junior synonym of auropunctata: Brown, 1948d: 102.
Type Material
- Tetramorium auropunctatum Roger, 1863: Syntype, worker(s), queen(s), male(s), Cuba.
Unless otherwise noted the text for the remainder of this section is reported from the publication that includes the original description.
Description
Worker
Cuezzo et al.(2015) - (n=10): HL: 0.55-0.61; HW: 0.42-0.48; EL: 0.11-0.13; SL: 0.35-0.40; AD: 0.20-0.27; PSL: 0.13-0.20; WL: 0.51-0.55; PD: 0.10-0.12; PTL: 0.12-0.15; PPTL: 0.10-0.12; PTW: 0.10; PPTW: 0.15-0.17; CI: 0.91-0.93; OI: 0.20-0.21.
Color reddish-yellow to orange brown, variable. Frons between frontal carinae punctate, covered with irregular striae. Only 5-6 striae reach vertexal margin of head. Occipital margin of head with short, curved setae. One curved and long seta present at posterior end of frontal carinae. There are three long hairs on each frontal carina, arranged longitudinally and curved inwards. Antenna with 11 segments. Shallow antennal scrobe, with sculpture similar to rest of head. Preocular carina runs along ventral margin of scrobe. Disc of clypeus striate. Clypeal striae weakly developed, running longitudinally. Masticatory margin of mandible with five teeth, no denticles and basal margin without teeth or denticles. Compound eye well developed, protruding from lateral margin of headin full face view. Malar space with 4-5 longitudinal irregular carinae. Vertexal margin straight. Promesonotum with 3-4 pairs of long, simple, and curved setae (length approx. 0.1 mm). Humeral angle well developed, without hairs. Mesosomal dorsum reticulate-punctuate. Propodeum with one pair of straight setae shorter than promesonotal setae. Long propodeal spines weakly curved inwards in dorsal view. In lateral view, propodeal spines shorter than or equal to length of petiole and posteriorly directed. Petiole rectangular, with 1-2 pairs of setae, similar in longitude to those of propodeum, anterior face well differentiated, forming a well defined angle with dorsal face. Mesosoma, petiole, and postpetiole, in lateral view, strongly spotted. Metapleural gland strongly developed, bulky. Posteropropodeal lobe rounded and well developed. Petiolar peduncle approximately of same length as petiole in lateral view. Very short acute spine present in anterior ventral face of peduncle. In dorsal view, petiole is long with rounded anterior edge, tapering towards apex. Postpetiole quadrate and wider than long, in dorsal view. Gaster weakly punctuate, with long, curved setae, scattered along each segment.
Queen
Cuezzo et al. (2015) - (n = 17): HL: 0.69-0.74; HW: 0.77-0.84; EL: 0.21-0.26; WL: 1.47-1.57.
Color and pilosity similar to worker. Head wider behind compound eyes. Scape barely reaches vertexal margin. Frontal carina separated by 10-12 longitudinal, poorly developed and irregular striae. Antennal scrobe deep and reticulate, with longitudinal carina that starts atventral margin of antennal torulus and almost reaches posterior margin of compound eye. Preocular carina runs along ventral margin of scrobe almost to occipital angle of head. Compound eye well-developed, located close to anterior margin of head, and protruding. Antena with 11 segments; apical club with two antennomeres. Mandibular dorsum with longitudinal, thin striae. Masticatory margin of mandible with five teeth. Malar space with three to four longitudinal striae. Disc of clypeus with more than 10 well developed, longitudinal striae. Pronotum poorly developed anterodorsally scutum encompasses more than half of mesosoma in dorsal view. Humeral angle rounded. Mesonotum dorsally striate, with thin, irregular striae and poorly developed. Axilla well developed, almost triangular and continuous in midline by narrow strip of integument. Middle area of scutellum with a deep, longitudinal groove. Anapleural sulcus completely divides anepisternum from the katepisternum. Anepisternum, katepisternum and metakatepisternum finely striate. Propodeal spine thick at its base. Propodeal lobe rounded. Fore and hind wings infuscate. Fore wing with three closed cells: costal, radial (=basal) and cubital (=subbasal). No closed discal cell. Hind wing with a closed radial cell and cubital cell, almost entirely delimited by veins. Hamuli composed by five hooks. Cinctus 1 and 2 well-developed. Petiolar peduncle longer than petiolar node in lateral view. Sterno-postpetiolar process sharp and short. Metasoma III weakly punctuate, covered with sparse, subdecumbent and thin setae.
Male
Cuezzo et al. (2015) - (n = 10): HL: 0.6-0.68; HW: 0.55-0.62; EL: 0.27-0.32; WL: 1.32-1.57.
Head oval in full face view. Mandible falcate with four teeth. Antenna with 13 segments, last antennomere of funicle longer than the rest. Scape not reaching frontovertexal margin of head. Compound eye well developed, covering more than half of lateral side of head. Three well developed ocelli are present; lateral ocellus reaches frontovertexal margin of head. Area between ocellicarinulate. Several diagonal carinae present in malar space and clypeal disc. Clypeal disc covered with parallel longitudinal carinae. Axillae medially compressed on middle of mesonotum; anterior and posterior margins nearly parallel. Fore and hind wing venation similar to queen. Petiolar node quadrate in profile, but with angles (anterior and posterior) not as strongly marked as worker. Postpetiole narrowly attached to abdominal segment III. Abdominal sternum IX (=subgenital plate) medially projected as a triangle and distally rounded. Pygostyles well developed, one segmented. Telomere elongated, finger like, ventrally curved, longer than basimere, distal width 0.05 mm. Digitus and cuspis poorly developed. Digitus stout and strongly curved ventrally. Ventral margin of aedeagus denticulate.
Gynandromorphs
Cuezzo et al. (2015) - (n = 2): HL: 0.70-0.75; HW: 0.70 (measured only in the specimen with a head shape similar to a normal male); EL: 0.32; SL (right scape): 0.50, (left scape): 0.25; WL: 1.60-1.62.
These are unusual sexual caste specimens because the left side of their head and antennae are similar to those of a normal male, but the right side has the characteristics of a queen. In one specimen, both compound eyes are less developed than in a normal male, but larger than a queen´s eyes (EL: 0.325). In the second specimen the left side of the head, including eyes and antennae, is similar to a male, while the right side is similar to a queen. In this specimen both antenna scrobes are well developed. In this last specimen, the left antenna is similar to a male and the right one has the same shape of the queen antenna. In both specimens, meso and metasoma are similar to a normal male, with similar wing venation and a male external genitalia. The only difference noted in the genitalia is referred to the development of the telomere. In both gynandromorphs the telomere is more stout (distal width: 0.1 mm) than in a normal male. Pygostyle is also longer than in a normal male.
Karyotype
- See additional details at the Ant Chromosome Database.
- Explore: Show all Karyotype data or Search these data. See also a list of all data tables or learn how data is managed.
- 2n = 32, karyotype = 20M+12A (Brazil) (deSouza et al., 2011).
- 2n = 32, karyotype = 16M+13SM+5ST (French Guiana) (Aguiar et al., 2020).
Worker Morphology
- Explore: Show all Worker Morphology data or Search these data. See also a list of all data tables or learn how data is managed.
- Caste: monomorphic
References
- Aguiar, H.J.A.C., Barros, L.A.C., Silveira, L.I., Petitclerc, F., Etienne, S., Orivel, J. 2020. Cytogenetic data for sixteen ant species from North-eastern Amazonia with phylogenetic insights into three subfamilies. Comparative Cytogenetics 14(1): 43–60 (doi:10.3897/CompCytogen.v14i1.46692).
- Ahuatzin, D.A., González-Tokman, D., Valenzuela-González, J.E., Escobar, F., Ribeiro, M.C., Acosta, J.C.L., Dáttilo, W. 2021. Sampling bias in multiscale ant diversity responses to landscape composition in a human-disturbed rainforest. Insectes Sociaux (doi:10.1007/s00040-021-00844-2).
- Albuquerque, E., Prado, L., Andrade-Silva, J., Siqueira, E., Sampaio, K., Alves, D., Brandão, C., Andrade, P., Feitosa, R., Koch, E., Delabie, J., Fernandes, I., Baccaro, F., Souza, J., Almeida, R., Silva, R. 2021. Ants of the State of Pará, Brazil: a historical and comprehensive dataset of a key biodiversity hotspot in the Amazon Basin. Zootaxa 5001, 1–83 (doi:10.11646/zootaxa.5001.1.1).
- Apolinário, L.da C.M.H., Almeida, Â.A.de, Queiroz, J.M., Vargas, A.B., Almeida, F.S. 2019. Diversity and guilds of ants in different land-use Systems in Rio de Janeiro State, Brazil. Floresta e Ambiente 26, e20171152 (doi:10.1590/2179-8087.115217).
- Baena, M.L., Escobar, F., Valenzuela, J.E. 2019. Diversity snapshot of green–gray space ants in two Mexican cities. International Journal of Tropical Insect Science 40, 239–250 (doi:10.1007/s42690-019-00073-y).
- Baker, A.J., Heraty, J.M., Mottern, J., Hang, J.Z., Hines, H.M., Lemmon, A.R., Lemmon, E.M. 2019. Inverse dispersal patterns in a group of ant parasitoids (Hymenoptera: Eucharitidae: Oraseminae) and their ant hosts. Systematic Entomology 45: 1–19 (doi:10.1111/syen.12371).
- Barros, L.A.C., Rabeling, C., Teixeira, G.A., dos Santos Ferreira Mariano, C., Delabie, J. H. C., de Aguiar, H. J. A. C. 2022. Decay of homologous chromosome pairs and discovery of males in the thelytokous fungus-growing ant Mycocepurus smithii. Scientific Reports 12, 4860 (doi:10.1038/s41598-022-08537-x).
- Bertelsmeier, C., A. Avril, O. Blight, A. Confais, L. Diez, H. Jourdan, J. Orivel, N. St Germes, and F. Courchamp. 2015a. Different behavioural strategies among seven highly invasive ant species. Biological Invasions. 17:2491-2503. doi:10.1007/s10530-015-0892-5
- Bertelsmeier, C., A. Avril, O. Blight, H. Jourdan, and F. Courchamp. 2015b. Discovery-dominance trade-off among widespread invasive ant species. Ecology and Evolution. 5:2673-2683. doi:10.1002/ece3.1542
- Borowiec, L. 2014. Catalogue of ants of Europe, the Mediterranean Basin and adjacent regions (Hymenoptera: Formicidae). Genus (Wroclaw) 25(1-2): 1-340.
- Borowiec, M.L., Moreau, C.S., Rabeling, C. 2020. Ants: Phylogeny and Classification. In: C. Starr (ed.), Encyclopedia of Social Insects (doi:10.1007/978-3-319-90306-4_155-1).
- Branstetter, M.G., Danforth, B.N., Pitts, J.P., Faircloth, B.C., Ward, P.S., Buffington, M.L., Gates, M.W., Kula, R.R., Brady, S.G. 2017. Phylogenomic insights into the evolution of stinging wasps and the origins of ants and bees. Current Biology 27, 1019–1025 (doi:10.1016/j.cub.2017.03.027).
- Brown, W. L., Jr. 1948d. The status of the genus Hercynia J. Enzmann (Hymenoptera: Formicidae). Entomol. News 59: 102 (page 102, Senior synonym of panamana)
- Bulter, I. 2020. Hybridization in ants. Ph.D. thesis, Rockefeller University.
- Calcaterra, L.A., Coulin, C., Briano, J.A., Follett, P.A. 2012. Acute exposure to low-dose radiation disrupts reproduction and shortens survival of Wasmannia auropunctata (Hymenoptera: Formicidae) queens. J Econ Entomol. 105(3): 817–822.
- Cardoso, D.C., Schoereder, J.H. 2014. Biotic and abiotic factors shaping ant (Hymenoptera: Formicidae) assemblages in Brazilian coastal sand dunes: The case of restinga in Santa Catarina. Florida Entomologist 97, 1443–1450. (doi:10.1653/024.097.0419).
- Chernyshova, A.M. 2021. A genetic perspective on social insect castes: A synthetic review and empirical study. M.S. thesis, The University of Western Ontario. Electronic Thesis and Dissertation Repository 7771.
- Chifflet, L., N. V. Guzman, O. Rey, V. A. Confalonieri, and L. A. Calcaterra. 2018. Southern expansion of the invasive ant Wasmannia auropunctata within its native range and its relation with clonality and human activity. Plos One. 13:16. doi:10.1371/journal.pone.0206602
- Conceição-Neto, R., França, E.C.B., Feitosa, R.M., Queiroz, J.M. 2021. Revisiting the ideas of trees as templates and the competition paradigm in pairwise analyses of ground-dwelling ant species occurrences in a tropical forest. Revista Brasileira de Entomologia 65, e20200026 (doi:10.1590/1806-9665-rbent-2020-0026).
- Correia, J.P.S.O., Mariano, C S F., Delabie, J.H.C., Lacau, S., Costa, M.A. 2016. Cytogenetic analysis of Pseudoponera stigma and Pseudoponera gilberti (Hymenoptera: Formicidae: Ponerinae): a taxonomic approach. Florida Entomologist 99: 718-721.
- Cuezzo, F., Calcaterra, L.A., Chifflet, L. and Follett, P. 2015. Wasmannia Forel (Hymenoptera: Formicidae: Myrmicinae) in Argentina: Systematics and distribution. Sociobiology. 62:246-265. doi:10.13102/sociobiology.v62i2.246-265
- Dekoninck, W., Wauters, N., Delsinne, T. 2019. Capitulo 35. Hormigas invasoras en Colombia. Hormigas de Colombia.
- Del Toro, I., Robbons, R.R., Pelini, S.L. 2012. The little things that run the world revisited: a review of ant-mediated ecosystem services and disservices (Hymenoptera: Formicidae). Myrmecological News 17: 133-146.
- Demetriou, J., Georgiadis, C., Roy, H., Martinou, A., Borowiec, L., Salata, S. 2022. One of the world’s worst invasive alien species Wasmannia auropunctata (Hymenoptera: Formicidae) detected in Cyprus. Sociobiology 69(4), e8536 (doi:10.13102/sociobiology.v69i4.8536).
- Devenish, A.J.M., Newton, R.J., Bridle, J.R., Gomez, C., Midgley, J.J., Sumner, S. 2021. Contrasting responses of native ant communities to invasion by an ant invader, Linepithema humile. Biological Invasions 23, 2553–2571 (doi:10.1007/s10530-021-02522-7).
- Deyrup, M., Davis, L. & Cover, S. 2000. Exotic ants in Florida. Transactions of the American Entomological Society 126, 293-325.
- Deyrup, M.A., Carlin, N., Trager, J., Umphrey, G. 1988. A review of the ants of the Florida Keys. Florida Entomologist 71: 163-176.
- do Nascimento, L.E., Amaral, R.R., Ferreira, R.M.dos A., Trindade, D.V.S., do Nascimento, R.E., da Costa, T.S., Souto, R.N.P. 2020. Ants (Hymenoptera: Formicidae) as potential mechanical vectors of pathogenic bacteria in a public hospital in the Eastern Amazon, Brazil. Journal of Medical Entomology 57: 1619–1626. (doi:10.1093/JME/TJAA062).
- Dolezal, A.G. 2019. Caste determination in arthropods. In: Encyclopedia of Animal Behavior, 2nd edition, Volume 4: 691–698 (doi:10.1016/B978-0-12-809633-8.20815-7).
- Espadaler, X., Pradero, C., Santana, J.A. 2018. The first outdoor-nesting population of Wasmannia auropunctata in continental Europe (Hymenoptera, Formicidae). Iberomyrmex 10: 1-8.
- Eyer, P.-A., Blumenfeld, A.J., Vargo, E.L. 2019. Sexually antagonistic selection promotes genetic divergence between males and females in an ant. Proceedings of the National Academy of Sciences 116, 24157–24163 (doi:10.1073/PNAS.1906568116).
- Fasi, J., Brodie, G. & Vanderwoude, C. 2013. Increases in crop pests caused by Wasmannia auropunctata in Solomon Islands subsistence gardens. Journal of Applied Entomology (doi: 10.1111/jen.12033).
- Fontenla, J.L., Brito, Y.M. 2011. Hormigas invasoras y vagabundas de Cuba. Fitosanidad 15(4), 253-259.
- Forel, A. 1886c. Diagnoses provisoires de quelques espèces nouvelles de fourmis de Madagascar, récoltées par M. Grandidier. Ann. Soc. Entomol. Belg. 30:ci-cvii. (page xlix, Combination in Ochetomyrmex)
- Forel, A. 1893j. Formicides de l'Antille St. Vincent, récoltées par Mons. H. H. Smith. Trans. Entomol. Soc. Lond. 1893: 333-418 (page 383, Combination in Wasmannia)
- Fournier, D., Estoup, A., Orivel, J., Foucaud, J., Jourdan, H., Le Breton, J., Keller, L. 2005. Clonal reproduction by males and females in the little fire ant. Nature (London) 435(7046): 1230-1234 (doi:10.1038/nature03705).
- Fournier, D., Tindo, M., Kenne, M., Mbenoun Masse, P.S., Van Bossche, V., De Coninck, E., Aron, S. 2012. Genetic structure, nestmate recognition and behaviour of two cryptic species of the invasive Big-Headed Ant Pheidole megacephala. PLoS ONE 7(2): e31480 (doi:10.1371/journal.pone.0031480).
- Franco, W., Ladino, N., Delabie, J.H.C., Dejean, A., Orivel, J., Fichaux, M., Groc, S., Leponce, M., Feitosa, R.M. 2019. First checklist of the ants (Hymenoptera: Formicidae) of French Guiana. Zootaxa 4674, 509–543 (doi:10.11646/zootaxa.4674.5.2).
- García-Padrón, L.Y., Amador, G.L., Delgado, M.M., Serrano, Y.M. 2020. Trophic ecology and morphology of Anolis bartschi (Squamata: Dactyloidae) in Viñales National Park, Cuba. Phyllomedusa: Journal of Herpetology 19, 177–187 (doi:10.11606/issn.2316-9079.v19i2p177-187).
- Glaser, S.M., Feitosa, R.M., Koch, A., Goß, N., do Nascimento, F.S., Grüter, C. 2021. Tandem communication improves ant foraging success in a highly competitive tropical habitat. Insectes Sociaux. (doi:10.1007/s00040-021-00810-y).
- Heraty, J.M., Rogers, D.V., Johnson, M.T., Perreira, W.D., Baker, A.J., Bitume, E., Murray, E., Varone, L. 2021. New record in the Hawaiian Islands of Orasema minutissima (Hymenoptera: Eucharitidae), an ant-parasitic wasp and a potential biocontrol agent against the Little Fire Ant, Wasmannia auropunctata (Hymenoptera: Formicidae). Bishop Museum Occasional Papers 137: 7–18.
- Herrera, H.W., Baert, L., Dekoninck, W., Causton, C.E., Sevilla, C.R., Pozo, P., Hendrickx, F. 2020. Distribution and habitat preferences of Galápagos ants (Hymenoptera: Formicidae). Belgian Journal of Entomology, 93: 1–60.
- Hoffmann, B., Eldridge, J., Marston, C. 2023. The first eradication of an exotic ant species from the entirety of Australia: Pheidole fervens. Management of Biological Invasions, 14(4), 619–624 (doi:10.3391/mbi.2023.14.4.03).
- Hornfeldt, J.A., Ohyama, L., Lucky, A. 2020. Using behavioral experimentation to understand the social structure of the Little Fire Ant (Wasmannia auropunctata) in Florida. University of Florida Journal of Undergraduate Research 22: 1-9.
- Jansen, G., Savolainen, R. 2010. Molecular phylogeny of the ant tribe Myrmicini (Hymenoptera: Formicidae). Zoological Journal of the Linnean Society 160(3), 482–495 (doi:10.1111/j.1096-3642.2009.00604.x).
- Jourdan, H., Sadlier, R.A., Bauer, A.M. 2000. Premières observations sur les conséquences de l’invasion de Wasmannia auropunctata 1863 (Roger) sur les prédateurs supérieurs dans les écosystèmes Néo-calédoniens. Actes des Collectes Insectes Sociaux 13, 121-126.
- Jourdan, H., Sadlier, R.A., Bauer, A.M. 2001. Little Fire Ant Invasion (Wasmannia auropunctata) as a threat to New Caledonian lizards: Evidences from a sclerophyll forest (Hymenoptera: Formicidae). Sociobiology 38, 283-301.
- Kaspari, M., Pickering, J., Longino, J., Windsor, D. 2001. The phenology of a Neotropical ant assemblage: evidence for continuous and overlapping reproduction. Behavioral Ecology and Sociobiology 50, 382–390 (doi:10.1007/s002650100378).
- Kempf, W. W. 1964e. Miscellaneous studies on Neotropical ants. III. (Hymenoptera: Formicidae). Stud. Entomol. 7: 45-71 (page 66, Senior synonym of glabra)
- Kirschenbaum, R. & Grace, J.K. 2008. Agonistic Responses of the Tramp Ants Anoplolepis gracilipes, Pheidole megacephala, Linepithema humile, and Wasmannia auropunctata (Hymenoptera: Formicidae). Sociobiology 51, 673-683.
- Koch, E.B.de A., Marques, T.E.D., Mariano, C.S.F., Neto, E.A.S., Arnhold, A., Peronti, A.L.B.G., Delabie, J.H.C. 2020. Diversity and structure preferences for ant-hemipteran mutualisms in cocoa trees (Theobroma cacao L., Sterculiaceae). Boletim do Museu Paraense Emílio Goeldi - Ciências Naturais 15, 65–81 (doi:10.46357/bcnaturais.v15i1.251).
- Ladino, N., Feitosa, R.M. 2022. Ants (Hymenoptera: Formicidae) of the Parque Estadual São Camilo, an isolated Atlantic Forest remnant in western Paraná, Brazil. ZOOLOGIA 39: e22001 (doi:10.1590/S1984-4689.v39.e22001).
- Landero-Torres, I., Garcia-Martinez, M.A., Galindo-Tovar, M.E., Leyva-Ovalle, O.R., Lee-Espinosa, H.E., Murguia-Gonzalez, J., Negrin-Ruiz, J. 2014. Alpha diversity of the myrmecofauna of the Natural Protected Area Metlac from Fortin, Veracruz, Mexico. Southwestern Entomologist 39: 541-553.
- Lau, M.K., Ellison, A.M., Nguyen, A., Penick, C., DeMarco, B., Gotelli, N.J., Sanders, N.J., Dunn, R.R., Helms Cahan, S. 2019. Draft Aphaenogaster genomes expand our view of ant genome size variation across climate gradients. PeerJ 7, e6447 (doi:10.7717/PEERJ.6447).
- Lee, C.-C., Weng, Y.-M., Lai, L.-C., Suarez, A.V., Wu, W.-J., Lin, C.-C., Yang, C.-C.S. 2020. Analysis of recent interception records reveals frequent transport of arboreal ants and potential predictors for ant invasion in Taiwan. Insects 11, 356 (doi:10.3390/INSECTS11060356).
- Lenoir, A., Chalon, Q., Carvajal, A., Ruel, C., Barroso, Á., Lackner, T., Boulay, R. 2012. Chemical integration of myrmecophilous guests in Aphaenogaster ant nests. Psyche: A Journal of Entomology 2012, 1–12 (doi:10.1155/2012/840860).
- Lopes, L.L., Mariano, C.S.F., Delabie, J.H.C., Silva, J.G. 2022. First cytogenetic study through conventional staining of the ant genus Blepharidatta Wheeler, 1915 (Hymenoptera: Formicidae: Attini). Sociobiology 69(4), e7843 (doi:10.13102/sociobiology.v69i4.7843).
- López-Bedoya, P.A., Gómez-Gaviria, M., Salazar-Fillippo, A.A., Pérez-Pedraza, L.F., Gutierrez-Cárdenas, P.D.A. 2023. Diet of Engystomops pustulosus (Anura: Leptodactylidae) from Colombia and current knowledge of its dietary ecology. Phyllomedusa 22(2): 159–170 (doi:10.11606/issn.2316-9079.v22i2p159-170).
- Lubertazzi, D. 2019. The ants of Hispaniola. Bulletin of the Museum of Comparative Zoology, 162(2), 59-210 (doi:10.3099/mcz-43.1).
- Lutinski, J., de Filtro, M., Baucke, L., Dorneles, F., Lutinski, C., Guarda, C. 2021. Ant assemblages (Hymenoptera: Formicidae) from areas under the direct influence of two small hydropower plants in Brazil. Brazilian Journal of Environmental Sciences (Online), 1-9 (doi:10.5327/Z217694781030).
- Mbenoun Masse, P. S., M. Tindo, M. Kenne, Z. Tadu, R. Mony, and C. Djieto-Lordon. 2017. Impact of the invasive ant Wasmannia auropunctata (Formicidae: Myrmicinae) on local ant diversity in southern Cameroon. African Journal of Ecology. 55:423-432. doi:10.1111/aje.12366
- Melo, T.S., Koch, E.B.A., Andrade, A.R.S., Travassos, M.L.O., Peres, M.C.L., Delabie, J.H.C. 2021. Ants (Hymenoptera: Formicidae) in different green areas in the metropolitan region of Salvador, Bahia state, Brazil. Brazilian Journal of Biology 82, e236269 (doi:10.1590/1519-6984.236269).
- Meurgey, F. 2020. Challenging the Wallacean shortfall: A total assessment of insect diversity on Guadeloupe (French West Indies), a checklist and bibliography. Insecta Mundi 786: 1–183.
- Mikheyev, A.S. & Mueller, U.G. 2007. Genetic relationships between native and introduced populations of the little fire ant Wasmannia auropunctata. Diversity and Distributions, 13, 573–579.
- Molfini, M., Zapparoli, M., Genovesi, P., Carnevali, L., Audisio, P., Di Giulio, A., Bologna, M.A. 2020. A preliminary prioritized list of Italian alien terrestrial invertebrate species. Biological Invasions 22(8), 2385–2399 (doi:10.1007/s10530-020-02274-w).
- Montgomery, M.P., Vanderwoude, C., Lynch, A.J.J., Robinson, W.A. 2020. The effects of laboratory rearing diet on recruitment behavior of Wasmannia auropunctata (Hymenoptera: Formicidaea). Florida Entomologist 103, 103-111 (doi:10.1653/024.103.0417).
- Moura, M.N., Cardoso, D.C., Cristiano, M.P. 2020. The tight genome size of ants: diversity and evolution under ancestral state reconstruction and base composition. Zoological Journal of the Linnean Society, zlaa135 (doi:10.1093/zoolinnean/zlaa135).
- Narváez-Vásquez, A., Gaviria, J., Vergara-Navarro, E.V., Rivera-Pedroza, L., Löhr, B. 2021. Ant (Hymenoptera: Formicidae) species diversity in secondary forest and three agricultural land uses of the Colombian Pacific Coast. Revista Chilena de Entomologia 47, 441–458 (doi:10.35249/rche.47.3.21.01).
- Nielsen, A., Atchison, R., Lucky, A. 2020. Effects of the invasive Little Fire Ant (Wasmannia auropunctata) on ant community composition on UF Campus. University of Florida | Journal of Undergraduate Research | Volume 22
- Oi, D.H., Lucky, A., Liebowitz, D.M. 2022. Response of Wasmannia auropunctata (Hymenoptera: Formicidae) to water-soaked Imported Fire Ant baits. Florida Entomologist 105(2): 108-114 (doi:10.1653/024.105.0202).
- Park, J., Xi, H., Park, J. 2021. Complete mitochondrial genome of the acrobat ant Crematogaster teranishii Santschi, 1930 (Formicidae; Hymenoptera). Mitochondrial DNA Part B 6, 593–595 (doi:10.1080/23802359.2021.1875922).
- Longino, J.T. & Fernández, F. 2007. Taxonomic review of the genus Wasmannia (pp. 271-289). In Snelling, R.R., Fisher, B.L. & Ward, P.S. (eds). Advances in ant systematics: homage to E.O. Wilson – 50 years of contributions. Memoirs of the American Entomological Institute 80: 690 pp. PDF
- Plentovich, S., Mizerek, T., Reeves, M.K., Amidon, F., Miller, S.E., Nanbara, M. 2020. Coastal strand and mangrove swamps of the Mariana Islands. Encyclopedia of the World’s Biomes, 2020, 185–197 (doi:10.1016/B978-0-12-409548-9.12422-4).
- Przybyszewski, K.R., Silva, R.J., Vicente, R.E., Garcia Freitas, J.V., Pereira, M.J.B., Izzo, T.J., Tonon, D.S. 2020. Can baited pitfall traps for sampling dung beetles replace conventional traps for sampling ants? Sociobiology 67, 376-387 (doi:10.13102/sociobiology.v67i3.5201).
- Rafael, J.A., Limeira-de-Oliveira, F., Hutchings, R.W., Miranda, G.F.G., Silva Neto, A.M.da, Somavilla, A., Camargo, A., Asenjo, A., Pinto, Â.P., Bello, A.de M., Dalmorra, C., Mello-Patiu, C.A.de, Carvalho, C.J.B.de, Takiya, D.M., Parizotto, D.R., Marques, D.W.A., Cavalheiro, D.de O., Mendes, D.M.de M., Zeppelini, D., Carneiro, E., Lima, É.F.B., Lima, E.C.A.de, Godoi, F.S.P.de, Pessoa, F.A.C., Vaz-de-Mello, F.Z., Sosa-Duque, F.J., Flores, H.F., Fernandes, I.O., Silva-Júnior, J.O., Gomes, L.R.P., Monné, M.L., Castro, M.C.M.de, Silva, M.P.G.da, Couri, M.S., Gottschalk, M.S., Soares, M.M.M., Monné, M.A., Rafael, M.S., Casagrande, M.M., Mielke, O.H.H., Grossi, P.C., Pinto, P.J.C., Bartholomay, P.R., Sobral, R., Heleodoro, R.A., Machado, R.J.P., Corrêa, R.C., Hutchings, R.S.G., Ale-Rocha, R., Santos, S.D.dos, Lima, S.P.de, Mahlmann, T., Silva, V.C., Fernandes, D.R.R. 2020. Insect (Hexapoda) diversity in the oceanic archipelago of Fernando de Noronha, Brazil: updated taxonomic checklist and new records. Revista Brasileira de Entomologia 64, e20200052 (doi:10.1590/1806-9665-rbent-2020-0052).
- Ramalho, M.de O., Kim, Z., Wang, S., Moreau, C.S. 2021. Wolbachia Across Social Insects: Patterns and Implications. Annals of the Entomological Society of America 114, 206–218 (doi:10.1093/aesa/saaa053).
- Ramos, A.S.de J.C., González, G., Lemos, R.N.S.de 2020. A new species of Diomus Mulsant (Coleoptera: Coccinellidae: Diomini) from the eastern Amazon. Revista Brasileira de Entomologia 64, e20200014 (doi:10.1590/1806-9665-rbent-2020-0014).
- Rey, O., Estoup, A., Vonshak, M., Loiseau, A., Blanchet, S., Calcaterra, L., Chifflet, L., Rossi, J.-P., Kergoat, G.J., Foucaud, J., Orivel, J., Leponce, M., Schultz, T. & Facon, B. 2012. Where do adaptive shifts occur during invasion? A multidisciplinary approach to unravelling cold adaptation in a tropical ant species invading the Mediterranean area. Ecology Letters 15, 1266–1275.
- Rey, O., Loiseau, A., Facon, B., Foucaud, J., Orivel, J., Cornuet, J.-M., Robert, S., Dobigny, G., Delabie, J.H.C., Mariano, C.D.S.F., Estoup, A. 2011. Meiotic recombination dramatically decreased in thelytokous aueens of the Little Fire Ant and their sexually produced workers. Molecular Biology and Evolution 28, 2591–2601 (doi:10.1093/molbev/msr082).
- Roger, J. 1863a. Die neu aufgeführten Gattungen und Arten meines Formiciden-Verzeichnisses nebst Ergänzung einiger früher gegebenen Beschreibungen. Berl. Entomol. Z. 7: 131-214 (page 182, worker, queen, male described)
- Rosas-Mejía, M., Guénard, B., Aguilar-Méndez, M. J., Ghilardi, A., Vásquez-Bolaños, M., Economo, E. P., Janda, M. 2021. Alien ants (Hymenoptera: Formicidae) in Mexico: the first database of records. Biological Invasions 23(6), 1669–1680 (doi:10.1007/s10530-020-02423-1).
- Rosumek, F. B. 2017. Natural History of Ants: What We (do not) Know about Trophic and Temporal Niches of Neotropical Species. Sociobiology. 64:244-255. doi:10.13102/sociobiology.v64i3.1623
- Siddiqui, J.A., Bamisile, B.S., Khan, M.M., Islam, W., Hafeez, M., Bodlah, I., Xu, Y. 2021. Impact of invasive ant species on native fauna across similar habitats under global environmental changes. Environmental Science and Pollution Research 28(39), 54362–54382 (doi:10.1007/s11356-021-15961-5).
- Silva, J. P., Valadares, L., Vieira, M. E. L., Teseo, S., Châline, N. 2021. Tandem running by foraging Pachycondyla striata workers in field conditions vary in response to food type, food distance, and environmental conditions. Current Zoology 67(5), 541–549 (doi:10.1093/cz/zoab050).
- Silva, P.S., Koch, E.B. de A., Arnhold, A., Delabie, J.H.C. 2022. Review of distribution modeling in ant (Hymenoptera: Formicidae) biogeographic studies. Sociobiology 69(4), e7775 (doi:10.13102/sociobiology.v69i4.7775).
- Smith, M. R. 1965. House-infesting ants of the eastern United States. Their recognition, biology, and economic importance. U. S. Dep. Agric. Tech. Bull. 1326: 1-105.
- Stukalyuk, S.V., Radchenko, A., Reshetov, A., Akhmedov, A., Goncharenko, I. 2021. Comparative analysis of the population structure of Crematogaster subdentata and Lasius neglectus in the primary and secondary ranges (Hymenoptera: Formicidae). Fragmenta Entomologica 53, 43-51 (doi:10.13133/2284-4880/436).
- Tibcherani, M., Aranda, R., Mello, R.L. 2020. Time to go home: The temporal threshold in the regeneration of the ant community in the Brazilian savanna. Applied Soil Ecology 150, 103451 (doi:10.1016/j.apsoil.2019.103451).
- Trible, W., McKenzie, S.K., Kronauer, D.J.C. 2020. Globally invasive populations of the clonal raider ant are derived from Bangladesh. Biology Letters 16, 20200105. (doi:10.1098/RSBL.2020.0105).
- Tseng, S.-P. 2020. Evolutionary history of a global invasive ant, Paratrechina longicornis (Dissertation_全文 ). Ph.D. thesis, Kyoto University.
- Ulysséa, M.A., Brandão, C.R.F. 2013. Ant species (Hymenoptera, Formicidae) from the seasonally dry tropical forest of northeastern Brazil: a compilation from field surveys in Bahia and literature records. Revista Brasileira de Entomologia 57, 217–224 (doi:10.1590/s0085-56262013005000002).
- Vandermeer, J., Perfecto, I. 2020. Endogenous spatial pattern formation from two intersecting ecological mechanisms: the dynamic coexistence of two noxious invasive ant species in Puerto Rico. Proceedings of the Royal Society B: Biological Sciences 287, 20202214 (doi:10.1098/rspb.2020.2214).
- Varela-Hernández, F., Medel-Zosayas, B., Martínez-Luque, E.O., Jones, R.W., De la Mora, A. 2020. Biodiversity in central Mexico: Assessment of ants in a convergent region. Southwestern Entomologist 454: 673-686.
- Wang, C., Sung, P.-J., Lin, C.-C., Ito, F., Billen, J. 2023. Parthenogenetic reproduction in Strumigenys ants: An update. Insects 14, 195 (doi:10.3390/insects14020195).
- Wetterer, J.K. 2013d. Worldwide spread of the little fire ant, Wasmannia auropunctata (Hymenoptera:Formicidae).Terrestrial Arthropod Reviews 6(2013) 173-184.
- Wetterer, J.K. 2017. Invasive ants of Bermuda revisited. Journal of Hymenoptera Research 54, 33–41 (doi:10.3897/jhr.54.11444).
- Wetterer, J.K. 2021. Ants (Hymenoptera, Formicidae) of St. Vincent, West Indies. Sociobiology 68, e6725 (doi:10.13102/sociobiology.v68i2.6725).
- Wetterer, J.K., Wetterer, A.L. 2004. Ants (Hymenoptera: Formicidae) of Bermuda. Florida Entomologist 87(2), 212–221 (doi:10.1653/0015-4040(2004)087[0212:ahfob2.0.CO;2]).
- Wheeler, G. C.; Wheeler, J. 1954d. The ant larvae of the myrmicine tribes Meranoplini, Ochetomyrmicini and Tetramoriini. Am. Midl. Nat. 52: 443-452 (page 444, larva described)
- Wheeler, W. M. 1922j. Ants of the American Museum Congo expedition. A contribution to the myrmecology of Africa. VIII. A synonymic list of the ants of the Ethiopian region. Bull. Am. Mus. Nat. Hist. 45: 711-1004 (page 912, Senior synonym of atomum)
References based on Global Ant Biodiversity Informatics
- Achury R., P. Chacon de Ulloa, and A. M. Arcila. 2008. Ant composition and competitive interactions with Wasmannia auropunctata in Tropical Dry Forest fragments. Revista Colombiana de Entomología 34 (2): 209-216.
- Achury R., and A.V. Suarez. 2017. Richness and composition of ground-dwelling ants in tropical rainforest and surrounding landscapes in the Colombian Inter-Andean valley. Neotropical Entomology https://doi.org/10.1007/s13744-017-0565-4
- Adams B. J., S. A. Schnitzer, and S. P. Yanoviak. 2016. Trees as islands: canopy ant species richness increases with the size of liana-free trees in a Neotropical forest. Ecography doi: 10.1111/ecog.02608
- Adams B. J., S. A. Schnitzer, and S. P. Yanoviak. 2019. Connectivity explains local ant community structure in a Neotropical forest canopy: a large-scale experimental approach. Ecology 100(6): e02673.
- Alonso L. E. 2010. A preliminary survey of the ants of the Kwamalasamutu region, SW Suriname. In: OShea, B.J., L.E. Alonso, & T.H. Larsen, (eds.). 2011. A Rapid Biological Assessment of the Kwamalasamutu region, Southwestern Suriname. RAP Bulletin of Biological Assessment 63. Conservation International, Arlington, VA.
- Alonso L. E., J. Persaud, and A. Williams. 2016. Biodiversity assessment survey of the south Rupununi Savannah, Guyana. BAT Survey Report No.1, 306 pages.
- Alvarez, G., I. Armbrecht, E. Jimenez, H. Armbrecht and P. Ulloa-Chacon. 2001. Ant-plant association in two Tococa species from a primary rain forest of Colombia Choco (Hymenoptera: Formicidae) Sociobiology 38(3B):585-602
- Amat-G G., M. G. Andrade-C. and F. Fernández. (eds.) 1999. Insectos de Colombia. Volumen II. Bogotá: Academia Colombiana de Ciencias Exactas, Físicas y Naturales, 433 pp. 131975
- Antoniazzi R., R. N. S. L. Garoo, W. Dattilo, S. P. Ribeiro, and F. S. Neves. 2019. Ant species richness and interactions in canopies of two distinct successional stages in a tropical dry forest. The Science of Nature 106: 20
- Araujo Castilho G., F. Barbosa Noll, E. R. da Silva, and E. F. dos Santos. 2011. Diversidade de Formicidae (Hymenoptera) em um fragmento de Floresta Estacional Semidecídua no Noroeste do estado de São Paulo, Brasil. R. bras. Bioci., Porto Alegre 9(2): 224-230.
- Armbrecht I., E. Jimenez, G. Alvarez, P. Ulloa-Chacon, and H. Armbrecht. 2001. An ant mosaic in the Colombian rain forest of Choco (Hymenoptera: formicidae0. Sociobiology 37(3B): 491-509.
- Besterlmeyer B. T., and R. R. Snelling. 2000. Ant (Hymenoptera: Formicidae) species recorded in Laguna de Tigre National Park, Petén, Guatemala. In: Brandon T. Bestelmeyer and Leeanne E. Alonso (eds.). 2000. A Biological Assessment of Laguna del Tigre National Park, Petén, Guatemala. RAP Bulletin of Biological Assessment 16, Conservation International, Washington, DC.
- Bicho C. L., M. L. C. Brancao, and S. M. Pires. 2007. Mirmecofauna (Hymenoptera, Formicidae) em hospitais e postos de Saude no municipio de Bagé, RS. Arq. Inst. Biol., São Paulo 74(4): 373-377.
- Boscardin J., E. C. Costa, J. H. C. Delabie. 2014. Relation between ant communities and environmental quality in Eucalyptus grandis submitted to different weedy species control in the south of Brazil. Entomotropica29(3): 173-182.
- Brandao, C.R.F. 1991. Adendos ao catalogo abreviado das formigas da regiao neotropical (Hymenoptera: Formicidae). Rev. Bras. Entomol. 35: 319-412.
- Brown W. L. Jr. 1948. The status of the genus Hercynia J. Enzmann (Hymenoptera: Formicidae). Entomological News 59: 102.
- Caldart V. M., S. Iop, J. A. Lutinski, and F. R. Mello Garcia. 2012. Ants diversity (Hymenoptera, Formicidae) of the urban perimeter of Chapecó county, Santa Catarina, Brazil. Revista Brasileira de Zoociências 14 (1, 2, 3): 81-94.
- Cardoso, D.C., T.G. Sobrinho and J.H. Schoereder. 2010. Ant community composition and its relationship with phytophysiognomies in a Brazilian Restinga. Insectes Sociaux 57:293-301
- Castano-Meneses G., R. De Jesus Santos, J. R. Mala Dos Santos, J. H. C. Delabie, L. L. Lopes, and C. F. Mariano. 2019. Invertebrates associated to Ponerine ants nests in two cocoa farming systems in the southeast of the state of Bahia, Brazil. Tropical Ecology 60: 52–61.
- Causton, C.E., C.R. Sevilla and S.D. Porter. 2005. Eradication of the Little Fire Ant, Wasmannia auropunctata (Hymenoptera: Formicidae), from Marchena Island, Galápagos: On the Edge of Success? Florida Entomologist 88(2):159-168
- Christianini A. V., A. J. Mayhé-Nunes, and P. S. Oliveira. 2012. Exploitation of Fallen Diaspores By Ants: Are There Ant-Plant Partner Choices? Biotropica 44: 360-367.
- Christianini A. V., and P. S. Oliveira. 2013. Edge effects decrease ant-derived benefits to seedlings in a neotropical savanna. Arthropod-Plant Interactions 7: 191-199.
- Clemes Cardoso D., and J. H. Schoereder. 2014. Biotic and abiotic factors shaping ant (Hymenoptera: Formicidae) assemblages in Brazilian coastal sand dunes: the case of restinga in Santa Catarina. Florida Entomologist 97(4): 1443-1450.
- Clemes Cardoso D., and M. Passos Cristiano. 2010. Myrmecofauna of the Southern Catarinense Restinga sandy coastal plain: new records of species occurrence for the state of Santa Catarina and Brazil. Sociobiology 55(1b): 229-239.
- Coelho M. S., G. W. Fernandes, J. C. Santos, and J. H. C. Delabie. 2009. Ants (Hymenoptera: Formicidae) as bioindicators of land restoration in a Brazilian Atlantic forest fragment. Sociobiology 54(1): 51-63.
- Correa M. M., W. D. Fernandes, and I. R. Leal. 2006. Ant Diversity (Hymenoptera: Formicidae) from Capões in Brazilian Pantanal: Relationship between Species Richness and Structural Complexity. Neotropical Entomology 35(6): 724-730.
- Costa-Milanez C. B., G. Lourenco-Silva, P. T. A. Castro, J. D. Majer, and S. P. Ribeiro. 2014. Are ant assemblages of Brazilian veredas characterised by location or habitat type? Braz. J. Biol. 74(1): 89-99.
- Cuezzo, F. 1998. Formicidae. Chapter 42 in Morrone J.J., and S. Coscaron (dirs) Biodiversidad de artropodos argentinos: una perspectiva biotaxonomica Ediciones Sur, La Plata. Pages 452-462.
- Dattilo W., N. Sibinel, J. C. F. Falcao, and R. V. Nunes. 2011. Ant fauna in a urban remnant of Atlantic forest in the municipuality of Marilia, state of Sao Paulo, Brazil. Bioscience Journal 27(3): 494-504.
- Davidson, D.W. 2005. Ecological stoichiometry of ants in a New World rain forest. Oecologia 142:221-231
- Delsinne T., Y. Roisin, and M. Leponce. 2007. Spatial and temporal foraging overlaps in a Chacoan ground-foraging ant assemblage. Journal of Arid Environments 71: 2944.
- Dias N. D. S., R. Zanetti, M. S. Santos, M. F. Gomes, V. Peñaflor, S. M. F. Broglio, and J. H. C. Delabie. 2012. The impact of coffee and pasture agriculture on predatory and omnivorous leaf-litter ants. Journal of Insect Science 13:29. Available online: http://www.insectscience.org/13.29
- Dias N. S., R. Zanetti, M. S. Santos, J. Louzada, and J. H. C. Delabie. 2008. Interaction between forest fragments and adjacent coffee and pasture agroecosystems: responses of the ant communities (Hymenoptera, Formicidae). Iheringia, Sér. Zool., Porto Alegre, 98(1): 136-142.
- Donoso D. A. 2014. Assembly mechanisms shaping tropical litter ant communities. Ecography 37 doi: 10.1111/j.1600-0587.2013.00253.x
- Donoso D. A., G. Onore, G. Ramon, and J. E. Lattke. 2014. Invasive ants of continental Ecuador, a first account. REMCB 35: 133-141.
- Drose W., L. R. Podgaiski, C. Fagundes Dias, M. de Souza Mendonca. 2019. Local and regional drivers of ant communities in forest-grassland ecotones in South Brazil: A taxonomic and phylogenetic approach. Plos ONE 14(4): e0215310.
- Emery C. 1894. Estudios sobre las hormigas de Costa Rica. Anales del Museo Nacional de Costa Rica 1888-1889: 45-64.
- Emery C. 1894. Studi sulle formiche della fauna neotropica. VI-XVI. Bullettino della Società Entomologica Italiana 26: 137-241.
- Emery C. 1906. Studi sulle formiche della fauna neotropica. XXVI. Bullettino della Società Entomologica Italiana 37: 107-194.
- Enzmann J. 1947. Hercynia, a new genus of myrmicine ants. J. N. Y. Entomol. Soc. 55: 43-46.
- Favretto M. A., E. Bortolon dos Santos, and C. J. Geuster. 2013. Entomofauna from West of Santa Catarina State, South of Brazil. EntomoBrasilis 6 (1): 42-63.
- Fernandes I., and J. de Souza. 2018. Dataset of long-term monitoring of ground-dwelling ants (Hymenoptera: Formicidae) in the influence areas of a hydroelectric power plant on the Madeira River in the Amazon Basin. Biodiversity Data Journal 6: e24375.
- Fernandes T. T., R. R. Silva, D. Rodrigues de Souza-Campana, O. Guilherme Morais da Silva, and M. Santina de Castro Morini. 2019. Winged ants (Hymenoptera: Formicidae) presence in twigs on the leaf litter of Atlantic Forest. Biota Neotropica 19(3): http://dx.doi.org/10.1590/1676-0611-bn-2018-0694
- Fernández F., E. E. Palacio, W. P. MacKay, and E. S. MacKay. 1996. Introducción al estudio de las hormigas (Hymenoptera: Formicidae) de Colombia. Pp. 349-412 in: Andrade M. G., G. Amat García, and F. Fernández. (eds.) 1996. Insectos de Colombia. Estudios escogidos. Bogotá: Academia Colombiana de Ciencias Exactas, Físicas y Naturales, 541 pp
- Fernández, F. and S. Sendoya. 2004. Lista de las hormigas neotropicales. Biota Colombiana Volume 5, Number 1.
- Fichaux M., B. Bechade, J. Donald, A. Weyna, J. H. C. Delabie, J. Murienne, C. Baraloto, and J. Orivel. 2019. Habitats shape taxonomic and functional composition of Neotropical ant assemblages. Oecologia 189(2): 501-513.
- Fleck M. D., E. Bisognin Cantarelli, and F. Granzotto. 2015. Register of new species of ants (Hymenoptera: Formicidae) in Rio Grande do Sul state. Ciencia Florestal, Santa Maria 25(2): 491-499.
- Forel A. 1908. Ameisen aus Sao Paulo (Brasilien), Paraguay etc. gesammelt von Prof. Herm. v. Ihering, Dr. Lutz, Dr. Fiebrig, etc. Verhandlungen der Kaiserlich-Königlichen Zoologisch-Botanischen Gesellschaft in Wien 58: 340-418.
- Forel A. 1908. Catálogo systemático da collecção de formigas do Ceará. Boletim do Museu Rocha 1(1): 62-69.
- Forel A. 1908. Fourmis de Costa-Rica récoltées par M. Paul Biolley. Bulletin de la Société Vaudoise des Sciences Naturelles 44: 35-72.
- Forel A. 1912. Formicides néotropiques. Part IV. 3me sous-famille Myrmicinae Lep. (suite). Mémoires de la Société Entomologique de Belgique. 20: 1-32.
- Franco W., N. Ladino, J. H. C. Delabie, A. Dejean, J. Orivel, M. Fichaux, S. Groc, M. Leponce, and R. M. Feitosa. 2019. First checklist of the ants (Hymenoptera: Formicidae) of French Guiana. Zootaxa 4674(5): 509-543.
- Gibernau M., J. Orivel, J. H. C. Delabie, D. Barabe, and A. Dejean. 2007. An asymmetrical relationship between an arboreal ponerine ant and a trash-basket epiphyte (Araceae). Biological Journal of the Linnean Society 91: 341-346.
- Groc S., J. H. C. Delabie, F. Fernandez, F. Petitclerc, B. Corbara, M. Leponce, R. Cereghino, and A. Dejean. 2017. Litter-dwelling ants as bioindicators to gauge the sustainability of small arboreal monocultures embedded in the Amazonian rainforest. Ecological Indicators 82: 43-49.
- Groc S., J. Orivel, A. Dejean, J. Martin, M. Etienne, B. Corbara, and J. H. C. Delabie. 2009. Baseline study of the leaf-litter ant fauna in a French Guianese forest. Insect Conservation and Diversity 2: 183-193.
- Guerrero R. J., and C. E. Sarmiento. 2010. Distribución altitudinal de hormigas (Hymenoptera, Formicidae) en la vertiente noroccidental de la Sierra Nevada de Santa Marta (Colombia). Acta Zoologica mexicana (n.s.) 26(2): 279-302.
- INBio Collection (via Gbif)
- IZIKO South Africa Museum Collection
- Iop S., V. M. Caldart, J. A. Lutinski, and F. R. Mello Garcia. 2009. Formigas urbanas da cidade de Xanxerê, Santa Catarina, Brasil. Biotemas 22(2): 55-64.
- Jacquemin J., T. Drouet, T. Delsinne, Y. Roisin, and M. Leponce. 2012. Soil properties only weakly affect subterranean ant distribution at small spatial scales. Applied Soil Ecology 62: 163-169.
- Jaffe, Klaus and Lattke, John. 1994. Ant Fauna of the French and Venezuelan Islands in the Caribbean in Exotic Ants, editor D.F. Williams. 182-190.
- Kamura C. M., M. S. C. Morini, C. J. Figueiredo, O. C. Bueno, and A. E. C. Campos-Farinha. 2007. Ant communities (Hymenoptera: Formicidae) in an urban ecosystem near the Atlantic Rainforest. Braz. J. Biol. 67(4): 635-641.
- Kaspari M. 1996. Litter ant patchiness at the 1-m 2 scale: disturbance dynamics in three Neotropical forests. Oecologia 107: 265-273.
- Kempf W. W. 1961. A survey of the ants of the soil fauna in Surinam (Hymenoptera: Formicidae). Studia Entomologica 4: 481-524.
- Kempf W. W. 1964e. Miscellaneous studies on Neotropical ants. III. (Hymenoptera: Formicidae). Studia Entomologica 7: 45-71.
- Kempf, W.W. 1972. Catalago abreviado das formigas da regiao Neotropical (Hym. Formicidae) Studia Entomologica 15(1-4).
- Kusnezov N. 1952. El género Wasmannia en la Argentina (Hymenoptera, Formicidae). Acta Zoologica Lilloana 10: 173-182.
- Kusnezov N. 1953. La fauna mirmecológica de Bolivia. Folia Universitaria. Cochabamba 6: 211-229.
- Kusnezov N. 1978. Hormigas argentinas: clave para su identificación. Miscelánea. Instituto Miguel Lillo 61:1-147 + 28 pl.
- Lapola D. M., and H. G. Fowler. 2008. Questioning the implementation of habitat corridors: a case study in interior São Paulo using ants as bioindicators. Braz. J. Biol., 68(1): 11-20.
- Lapolla, J.S., T. Suman, J. Soso-Calvo and T.R. Schultz. 2006. Leaf litter ant diversity in Guyana. Biodiversity and Conservation 16:491510
- LeBrun, E.G., C. V. Tillberg, A. V. Suarez, P. J. Folgarait, C. R. Smith and D. A. Holway. 2007. An Experimental Study of Competition between Fire Ants and Argentine Ants in Their Native Range. Ecology 88(1):63-75
- Leponce M., J. H. C. Delabie, J. Orivel, J. Jacquemin, M. Calvo Martin, and A. Dejean. 2019. Tree-dwelling ant survey (Hymenoptera, Formicidae) in Mitaraka, French Guiana, in Touroult J. (ed.), “Our Planet Reviewed” 2015 large-scale biotic survey in Mitaraka, French Guiana. Zoosystema 41 (10): 163-179.
- Levings S. C. 1983. Seasonal, annual, and among-site variation in the ground ant community of a deciduous tropical forest: some causes of patchy species distributions. Ecological Monographs 53(4): 435-455.
- Longino J. T. L., and M. G. Branstetter. 2018. The truncated bell: an enigmatic but pervasive elevational diversity pattern in Middle American ants. Ecography 41: 1-12.
- Longino J. T. and Fernández, F. 2007. Taxonomic review of the genus Wasmannia. Memoirs of the American Entomological Institute 80: 271-289
- Longino J. T., and N. M. Nadkarni. 1990. A comparison of ground and canopy leaf litter ants (Hymenoptera: Formicidae) in a Neotropical montane forest. Psyche (Cambridge) 97: 81-94.
- Longino J. T., and R. K. Colwell. 2011. Density compensation, species composition, and richness of ants on a neotropical elevational gradient. Ecosphere 2(3): 16pp.
- Longino J. et al. ADMAC project. Accessed on March 24th 2017 at https://sites.google.com/site/admacsite/
- Lopes D. T., J. Lopes, I. Cardoso do Nascimento, and J. H. Delabie. 2010. Epigeic ants diversity (Hymenoptera, Formicidae) in three environments in Mata dos Godoy State Park, Londrina, State of Paraná, Brazil. Iheringia, Sér. Zool., Porto Alegre, 100(1): 84-90.
- Lopes M. C., G. P. A. Lamarre, C. Baraloto, P. V. A. Fine, A. Vincentini, and F. B. Baccaro. 2019. The Amazonas-trap: a new method for sampling plant-inhabiting arthropod communities in tropical forest understory. Entomologia Experimentalis et Applicata https://doi.org/10.1111/eea.12797
- Luederwaldt H. 1918. Notas myrmecologicas. Rev. Mus. Paul. 10: 29-64.
- Lutinski J. A., B. C. Lopes, and A. B. B.de Morais. 2013. Diversidade de formigas urbanas (Hymenoptera: Formicidae) de dez cidades do sul do Brasil. Biota Neotrop. 13(3): 332-342.
- Lutinski J. A., F. R. Mello Garcia, C. J. Lutinska, and S. Iop. 2008. Ants diversity in Floresta Nacional de Chapecó in Santa Catarina State, Brazil. Ciência Rural, Santa Maria 38(7): 1810-1816.
- Macedo-Reis L. E., A. C. Leite, T. J. Guerra, R. Antoniazzi, and F. Neves. Suspended leaf litter in an understorey treelet as habitat extension for ground-dwelling ants in the Atlantic Forest, south-eastern Brazil. Journal of Tropical Ecology https://doi.org/10.1017/S0266467419000154
- Maes, J.-M. and W.P. MacKay. 1993. Catalogo de las hormigas (Hymenoptera: Formicidae) de Nicaragua. Revista Nicaraguense de Entomologia 23.
- Mann W. M. 1916. The Stanford Expedition to Brazil, 1911, John C. Branner, Director. The ants of Brazil. Bulletin of the Museum of Comparative Zoology 60: 399-490
- Mann W. M. 1922. Ants from Honduras and Guatemala. Proceedings of the United States National Museum 61: 1-54.
- Marinho C. G. S., R. Zanetti, J. H. C. Delabie, M. N. Schlindwein, and L. de S. Ramos. 2002. Ant (Hymenoptera: Formicidae) Diversity in Eucalyptus (Myrtaceae) Plantations and Cerrado Litter in Minas Gerais, Brazil. Neotropical Entomology 31(2): 187-195.
- Medeiros Macedo L. P., E. B. Filho, amd J. H. C. Delabie. 2011. Epigean ant communities in Atlantic Forest remnants of São Paulo: a comparative study using the guild concept. Revista Brasileira de Entomologia 55(1): 7578.
- Menozzi C. 1931. Contribuzione alla conoscenza del microgenton di Costa Rica. III. Hymenoptera - Formicidae. Bollettino del Laboratorio di Zoologia Generale e Agraria della Reale Scuola Superiore d'Agricoltura. Portici. 25: 259-274.
- Mertl A. L., J. F. A. Traniello, K. Ryder Wilkie, and R. Constantino. 2012. Associations of two ecologically significant social insect taxa in the litter of an amazonian rainforest: is there a relationship between ant and termite species richness? Psyche doi:10.1155/2012/312054
- Morini M. S. de C., C. de B. Munhae, R. Leung, D. F. Candiani, and J. C. Voltolini. 2007. Comunidades de formigas (Hymenoptera, Formicidae) em fragmentos de Mata Atlântica situados em áreas urbanizadas. Iheringia, Sér. Zool., Porto Alegre, 97(3): 246-252.
- Nascimento Santos M., J. H. C. Delabie, and J. M. Queiroz. 2019. Biodiversity conservation in urban parks: a study of ground-dwelling ants (Hymenoptera: Formicidae) in Rio de Janeiro City. Urban Ecosystems https://doi.org/10.1007/s11252-019-00872-8
- Navarro, E.V. Vergara, H. Echavarria Sanchez, F.J. Serna Cardona. 2007. Hormigas (Hymenoptera: Formicidae) asociadas al arboretum de la Universidad Nacional de Colombia, sede Medellin. Boletín Sociedad Entomológica Aragonesa 40:497-505.
- Neves F. S., R. F. Braga, M. M. do Espirito-Santo, J. H. C. Delabie, G. Wilson Fernandes, and G. A. Sanchez-Azofeifa. 2010. Diversity of Arboreal Ants In a Brazilian Tropical Dry Forest: Effects Of Seasonality and Successional Stage. Sociobiology 56(1): 1-18.
- Nunes F. A., G. B. Martins Segundo, Y. B. Vasconcelos, R. Azevedo, and Y. Quinet. 2011. Ground-foraging ants (Hymenoptera: Formicidae) and rainfall effect on pitfall trapping in a deciduous thorn woodland (Caatinga), Northeastern Brazil. Rev. Biol. Trop. 59 (4): 1637-1650.
- Oliveira-Santos L. G. R., R. D. Loyola, A. B. Vargas. 2009. Canopy Traps: a Technique for Sampling Arboreal Ants in Forest Vertical Strata. Neotropical Entomology 38(5):691-694.
- Osorio Rosado J. L, M. G. de Goncalves, W. Drose, E. J. Ely e Silva, R. F. Kruger, and A. Enimar Loeck. 2013. Effect of climatic variables and vine crops on the epigeic ant fauna (Hymenoptera: Formicidae) in the Campanha region, state of Rio Grande do Sul, Brazil. J Insect Conserv 17: 1113-1123.
- Ottonetti L., L. Tucci, F. Frizzi, G. Chelazzi, and G. Santini. 2010. Changes in ground-foraging ant assemblages along a disturbance gradient in a tropical agricultural landscape. Ethology Ecology & Evolution 22: 7386.
- Pacheco R., H. L. Vasconcelos, S. Groc, G. P. Camacho, and T. L. M. Frizzo. 2013. The importance of remnants of natural vegetation for maintaining ant diversity in Brazilian agricultural landscapes. Biodivers. Conserv. DOI 10.1007/s10531-013-0463-y
- Pacheco R., and H. L. Vasconcelos. 2012. Subterranean Pitfall Traps: Is ItWorth Including Them in Your Ant Sampling Protocol? Psyche doi:10.1155/2012/870794
- Perfecto I., and J. Vandermeer. 1996. Microclimatic changes and the indirect loss of ant diversity in a tropical agroecosystem. Oecologia 108: 577-582.
- Pignalberi C. T. 1961. Contribución al conocimiento de los formícidos de la provincia de Santa Fé. Pp. 165-173 in: Comisión Investigación Científica; Consejo Nacional de Investigaciones Científicas y Técnicas (Argentina) 1961. Actas y trabajos del primer Congreso Sudamericano de Zoología (La Plata, 12-24 octubre 1959). Tomo III. Buenos Aires: Librart, 276 pp.
- Pires de Prado L., R. M. Feitosa, S. Pinzon Triana, J. A. Munoz Gutierrez, G. X. Rousseau, R. Alves Silva, G. M. Siqueira, C. L. Caldas dos Santos, F. Veras Silva, T. Sanches Ranzani da Silva, A. Casadei-Ferreira, R. Rosa da Silva, and J. Andrade-Silva. 2019. An overview of the ant fauna (Hymenoptera: Formicidae) of the state of Maranhao, Brazil. Pap. Avulsos Zool. 59: e20195938.
- Piva A., and A. E. de C. Campos. 2012. Ant Community Structure (Hymenoptera: Formicidae) in Two Neighborhoods with Different Urban Profiles in the City of Sao Paulo, Brazil. Psyche 2012 (390748): 1-8
- Ramos L. S., R. Z. B. Filho, J. H. C. Delabie, S. Lacau, M. F. S. dos Santos, I. C. do Nascimento, and C. G. S. Marinho. 2003. Ant communities (Hymenoptera: Formicidae) of the leaf-litter in cerrado stricto sensu areas in Minas Gerais, Brazil. Lundiana 4(2): 95-102.
- Ramos L. de S., C. G. S. Marinho, R. Zanetti, and J. H. C. Delabie. 2003. Impacto de iscas formicidas granuladas sobre a mirmecofauna não-alvo em eucaliptais segundo duas formas de aplicacação / Impact of formicid granulated baits on non-target ants in eucalyptus plantations according to two forms of application. Neotropical Entomology 32(2): 231-237.
- Ramos L. de S., R. Zanetti, C. G. S. Marinho, J. H. C. Delabie, M. N. Schlindwein, and R. P. Almado. 2004. Impact of mechanical and chemical weedings of Eucalyptus grandis undergrowth on an ant community (Hymenoptera: Formicidae). Rev. Árvore 28(1): 139-146.
- Resende J. J., G. M. de M. Santos, I. C. do Nascimento, J. H. C. Delabie, and E. M. da Silva. 2011. Communities of ants (Hymenoptera Formicidae) in different Atlantic rain forest phytophysionomies. Sociobiology 58(3): 779-799.
- Ribas C. R., F. A. Schmidt, R. R. C. Solar, R. B. F. Campos, C. L. Valentim, and J. H. Schoereder. 2012. Ants as Indicators of the Success of Rehabilitation Efforts in Deposits of Gold Mining Tailings. Restoration Ecology 20(6): 712722.
- Ribas C. R., J. H. Schoereder, M. Pic, and S. M. Soares. 2003. Tree heterogeneity, resource availability, and larger scale processes regulating arboreal ant species richness. Austral Ecology 28(3): 305-314.
- Rodriguez E. R., and J. E. Lattke. 2012. Diversidad de hormigas en un gradiente altitudinal de la cordillera de la Costa, Venezuela. Boletín de la Sociedad Entomológica Aragonesa (S.E.A.) 50: 295?304.
- Rosa da Silva R. 1999. Formigas (Hymenoptera: Formicidae) do oeste de Santa Catarina: historico das coletas e lista atualizada das especies do Estado de Santa Catarina. Biotemas 12(2): 75-100.
- Rosumek, F.B., M.A. Ulyssea, B.C. Lopes, J. Steiner. 2008. Formigas de solo e de bromélias em uma área de Mata Atlântica, Ilha de Santa Catarina, sul do Brasil: Levantamento de espécies e novos registros. Revista Biotemas 21(4):81-89.
- Salinas P. J. 2010. Catalogue of the ants of the Táchira State, Venezuela, with notes on their biodiversity, biogeography and ecology (Hymenoptera: Formicidae: Amblyioponinae, Ponerinae, Proceratiinae, Myrmicinae, Ecitoninae, Formicinae, Pseudomyrmecinae, Dolichoderinae). Boletín de la SEA 47: 315-328.
- Sandoval V. E., and G. Zambrano. 2007. Catálogo de las hormigas presentes en el Museo de Historia Natural de la Universidad del Cauca. Taller Editorial de la Universidad del Cauca, Popayán. 60 pp.
- Santoandre S., J. Filloy, G. A. Zurita, and M. I. Bellocq. 2019. Ant taxonomic and functional diversity show differential response to plantation age in two contrasting biomes. Forest Ecology and Management 437: 304-313.
- Santos Lopes J. F., N. Martins dos Reis Hallack, T. Archanjo de Sales, M. Silva Brugger, L. F. Ribeiro, I. N. Hastenreiter, and R. da Silva Camargo. 2012. Comparison of the Ant Assemblages in Three Phytophysionomies: Rocky Field, Secondary Forest, and Riparian ForestA Case Study in the State Park of Ibitipoca, Brazil. Psyche doi:10.1155/2012/928371
- Santos M. S., J. N. C. Louzada, N. Dias, R. Zanetti, J. H. C. Delabie, and I. C. Nascimento. 2006. Litter ants richness (Hymenoptera, Formicidae) in remnants of a semi-deciduous forest in the Atlantic rain forest, Alto do Rio Grande region, Minas Gerais, Brazil. Iheringia, Sér. Zool., Porto Alegre, 96(1): 95-101.
- Santschi F. 1925. Fourmis des provinces argentines de Santa Fe, Catamarca, Santa Cruz, Córdoba et Los Andes. Comunicaciones del Museo Nacional de Historia Natural "Bernardino Rivadavia" 2: 149-168.
- Santschi F. 1929. Nouvelles fourmis de la République Argentine et du Brésil. Anales de la Sociedad Cientifica Argentina. 107: 273-316.
- Santschi F. 1931. Contribution à l'étude des fourmis de l'Argentine. Anales de la Sociedad Cientifica Argentina. 112: 273-282.
- Santschi F. 1931. Fourmis de Cuba et de Panama. Revista de Entomologia (Rio de Janeiro). 1: 265-282.
- Schmidt F. A., and R. R. C. Solar. Is it important to collect hypogaeic ants? How to collect them? Biológico, São Paulo 69(2): 267-270.
- Schmidt K., R. Corbetta, and A.J. Aires de Carmargo. 2005. Formigas (Hymenoptera: Formicidae) da Ilha João da Cunha, SC: composição e diversidade. Biotemas, 18 (1): 57 - 71
- Schmidt, F.A. and R.R.C Solar. 2010. Hypogaeic pitfall traps: methodological advances and remarks to improve the sampling of a hidden ant fauna. Insectes Sociaux 57:261-266.
- Schoereder J. H., T. G. Sobrinho, M. S. Madureira, C. R. Ribas, and P. S. Oliveira. 2010. The arboreal ant community visiting extrafloral nectaries in the Neotropical cerrado savanna. Terrestrial Arthropod Reviews 3: 3-27.
- Silva F. H. O., J. H. C. Delabie, G. B. dos Santos, E. Meurer, and M. I. Marques. 2013. Mini-Winkler Extractor and Pitfall Trap as Complementary Methods to Sample Formicidae. Neotrop Entomol 42: 351358.
- Silva F. R., R. M. Begnini, V. A. Klier, K. Z. Scherer, B. C. Lopes, and T. T. Castellani. 2009. Syagrus romanzoffi ana (Arecaceae) Seed Utilization by Ants in a Secondary Forest in South Brazil. Neotropical Entomology 38(6): 873-875.
- Silva R. R., R. S. Machado Feitosa, and F. Eberhardt. 2007. Reduced ant diversity along a habitat regeneration gradient in the southern Brazilian Atlantic Forest. Forest Ecology and Management 240: 61-69.
- Silva R.R., and C. R. F. Brandao. 2014. Ecosystem-Wide Morphological Structure of Leaf-Litter Ant Communities along a Tropical Latitudinal Gradient. PLoSONE 9(3): e93049. doi:10.1371/journal.pone.0093049
- Silvestre R., C. R. F. Brandão, and R. R. Silva da 2003. Grupos funcionales de hormigas: el caso de los gremios del cerrado. Pp. 113-148 in: Fernández, F. (ed.) 2003. Introducción a las hormigas de la región Neotropical. Bogotá: Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, xxvi + 424 pp.
- Silvestre R., M. F. Demetrio, and J. H. C. Delabie. 2012. Community Structure of Leaf-Litter Ants in a Neotropical Dry Forest: A Biogeographic Approach to Explain Betadiversity. Psyche doi:10.1155/2012/306925
- Siqueira de Castro F., A. B. Gontijo, P. de Tarso Amorim Castro, and S. Pontes Ribeiro. 2012. Annual and Seasonal Changes in the Structure of Litter-Dwelling Ant Assemblages (Hymenoptera: Formicidae) in Atlantic Semideciduous Forests. Psyche doi:10.1155/2012/959715
- Siqueira de Castro F., A. B. Gontijo, W. Duarte da Rocha, and S. Pontes Ribeiro. 2011. As comunidades de formigas de serapilheira nas florestas semidecíduas do Parque Estadual do Rio Doce, Minas Gerais. MG.BIOTA, Belo Horizonte 3(5): 5-24.
- Smith M. A., W. Hallwachs, D. H. Janzen. 2014. Diversity and phylogenetic community structure of ants along a Costa Rican elevational gradient. Ecography 37(8): 720-731.
- Soares S. A., D. Lange, and W. F. Antoniali Junior. 2006. Communities of Epigaeic ants (Hymenoptera: Formicidae) in an area of reforestation and in native forest. Sociobiology 49(3): 251-263.
- Soares S. A., W. F. Antoniali Junior, and S. E. Lima-Junior. 2010. Diversidade de formigas epigéicas (Hymenoptera, Formicidae) em dois ambientes no Centro-Oeste do Brasil. Revista Brasileira de Entomologia 54(1): 7681.
- Sobrinho T. G., and J. H. Schoereder. 2007. Edge and shape effects on ant (Hymenoptera: Formicidae) species richness and composition in forest fragments. Biodivers Conserv 16: 14591470.
- Sobrinho T., J. H. Schoereder, C. F. Sperber, and M. S. Madureira. 2003. Does fragmentation alter species composition in ant communities (Hymenoptera: Formicidae)? Sociobiology 42(2): 329-342.
- Solomon, S.E. and A.S. Mikheyev. 2005. The ant (Hymenoptera: Formicidae) fauna of Cocos Island, Costa Rica. Florida Entomologist 88(4):415-423
- Sosa-Calvo J. 2007. Ants of the leaf litter of two plateaus in Eastern Suriname. In Alonso, L.E. and J.H. Mol (eds.). 2007. A rapid biological assessment of the Lely and Nassau plateaus, Suriname (with additional information on the Brownsberg Plateau). RAP Bulletin of Biological Assessment 43. Conservation International, Arlington, VA, USA.
- Suguituru S. S., M. Santina de Castro Morini, R. M. Feitosa, and R. Rosa da Silva. 2015. Formigas do Alto Tiete. Canal 6 Editora 458 pages
- Ulloa Chacon P., M. L. Baena, J. Bustos, R. C. Aldana, J. A. Aldana, and M. A. Gamboa. 1996. Fauna de hormigas del departamento del Valle del Cauca (Colombia). Pp. 413-451. In Andrade-C M. G., G. Amat Garcia, and F. Fernandez. Insectoss de Colombia, estudios escogidos.
- Ulyssea M. A., B. C. Lopes, A. Zillidens, J. Steiner. 2007. Ants associated to Nidularium innocentii and Aechmea lindenii (Bromeliaceae) in atlantic rainforest in southern Brazil. Biológico, São Paulo 69(2): 319-324.
- Ulyssea M. A., and C. R. F. Brandao. 2013. Ant species (Hymenoptera, Formicidae) from the seasonally dry tropical forest of northeastern Brazil: a compilation from field surveys in Bahia and literature records. Revista Brasileira de Entomologia 57(2): 217224.
- Ulyssea M.A., C. E. Cereto, F. B. Rosumek, R. R. Silva, and B. C. Lopes. 2011. Updated list of ant species (Hymenoptera, Formicidae) recorded in Santa Catarina State, southern Brazil, with a discussion of research advances and priorities. Revista Brasileira de Entomologia 55(4): 603-611.
- Ulysséa M. A., C. R. F. Brandão. 2013. Ant species (Hymenoptera, Formicidae) from the seasonally dry tropical forest of northeastern Brazil: a compilation from field surveys in Bahia and literature records. Revista Brasileira de Entomologia 57(2): 217-224.
- Vasconcelos, H.L., J.M.S. Vilhena, W.E. Magnusson and A.L.K.M. Albernaz. 2006. Long-term effects of forest fragmentation on Amazonian ant communities. Journal of Biogeography 33:1348-1356
- Viana-Silva F. E. C., and C. M. Jacobi. 2012. Myrmecofauna of Ironstone Outcrops: Composition and Diversity. Neotrop Entomol 41: 263271.
- Vittar, F. 2008. Hormigas (Hymenoptera: Formicidae) de la Mesopotamia Argentina. INSUGEO Miscelania 17(2):447-466
- Vittar, F., and F. Cuezzo. "Hormigas (Hymenoptera: Formicidae) de la provincia de Santa Fe, Argentina." Revista de la Sociedad Entomológica Argentina (versión On-line ISSN 1851-7471) 67, no. 1-2 (2008).
- Weber N. A. 1934. Notes on neotropical ants, including the descriptions of new forms. Revista de Entomologia (Rio de Janeiro) 4: 22-59.
- Weber N. A. 1938. The food of the giant toad, Bufo marinus (L.), in Trinidad and British Guiana with special reference to the ants. Annals of the Entomological Society of America 31: 499-503.
- Weber N. A. 1947. Lower Orinoco River fungus-growing ants (Hymenoptera: Formicidae, Attini). Boletín de Entomologia Venezolana 6: 143-161.
- Wetterer J. K., and S. D. Porter. 2003. The little fire ant, Wasmannia auropunctata: distribution, impact and control. Sociobiology 41(3): 1-41.
- Wheeler W. M. 1922. The ants of Trinidad. American Museum Novitates 45: 1-16.
- Wheeler W. M. 1925. Neotropical ants in the collections of the Royal Museum of Stockholm. Arkiv för Zoologi 17A(8): 1-55.
- Wheeler W. M. 1942. Studies of Neotropical ant-plants and their ants. Bulletin of the Museum of Comparative Zoology 90: 1-262.
- Wheeler, William Morton. 1916. Ants Collected in Trinidad by Professor Roland Thaxter, Mr. F. W. Urich, and Others. Bulletin of the Museum of Comparitive Zoology at Harvard University. 40(8):322-330
- Wild, A. L. "A catalogue of the ants of Paraguay (Hymenoptera: Formicidae)." Zootaxa 1622 (2007): 1-55.
- Yanoviak S. P., S, M. Berghoff, K. E. Linsenmair, and G. Zotz. 2011. Effects of an Epiphytic Orchid on Arboreal Ant Community Structure in Panama. Biotropica 43(6): 731-737.
- Yanoviak S. P., and M. Kaspari. 2000. Community structure and the habitat templet: ants in the tropical forest canopy and litter. Oikos 89: 259-266.
- Zolessi L. C. de, Y. P. Abenante, and M. E. de Philippi. 1988. Lista sistematica de las especies de Formicidos del Uruguay. Comun. Zool. Mus. Hist. Nat. Montev. 11: 1-9.
- Zolessi L. C. de; Y. P. de Abenante, and M. E. Philippi. 1989. Catálogo sistemático de las especies de Formícidos del Uruguay (Hymenoptera: Formicidae). Montevideo: ORCYT Unesco, 40 + ix pp.
- da Silva Araujo, M., Castro Della Lucia, T.M., DA VEIGA, Clayton E y CARDOSO DO NASCIMENTO, Ivan. 2004. Efeito da queima da palhada de cana-de-açúcar sobre comunidade de formicídeos. Ecol. austral. 14(2): 191-200.
- da Silva de Oliveira A. B., and F. A. Schmidt. 2019. Ant assemblages of Brazil nut trees Bertholletia excelsa in forest and pasture habitats in the Southwestern Brazilian Amazon. Biodiversity and Conservation 28(2): 329-344.
- da Silva, R.R., C.R.F. Brandao, and R. Silvestre. 2004. Similarity Between Cerrado Localities in Central and Southeastern Brazil Based on the Dry Season Bait Visitors Ant Fauna. Studies on Neotropical Fauna and Environment 39(3):191-199.
- de Zolessi, L.C., Y.P. de Abenante and M.E. Philippi. 1987. Lista sistemática de las especies de formícidos del Uruguay. Comunicaciones Zoologicas del Museo de Historia Natural de Montevideo 11(165):1-9
- de Zolessi, L.C., Y.P. de Abenante and M.E. Phillipi. 1989. Catalago Systematico de las Especies de Formicidos del Uruguay (Hymenoptera: Formicidae). Oficina Regional de Ciencia y Technologia de la Unesco para America Latina y el Caribe- ORCYT. Montevideo, Uruguay
- van der Hammen T., and P. S. Ward. 2005. Ants from the Ecoandes expeditions: diversity and distribution. Studies on Tropical Andean Ecosystems 6: 239-248.
- Pages using DynamicPageList3 parser function
- Common Name
- Highly invasive
- Parthenogenetic
- Supercolonies
- North subtropical
- Tropical
- South subtropical
- South temperate
- Eucharitid wasp Associate
- Host of Orasema minutissima
- FlightMonth
- Karyotype
- Species
- Extant species
- Formicidae
- Myrmicinae
- Attini
- Wasmannia
- Wasmannia auropunctata
- Myrmicinae species
- Attini species
- Wasmannia species
- Ssr