Tetramorium forte
| Tetramorium forte | |
|---|---|

| |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Arthropoda |
| Class: | Insecta |
| Order: | Hymenoptera |
| Family: | Formicidae |
| Subfamily: | Myrmicinae |
| Tribe: | Crematogastrini |
| Genus: | Tetramorium |
| Species: | T. forte |
| Binomial name | |
| Tetramorium forte Forel, 1904
| |
| Synonyms | |
| |


{kind=link}
A species that has had a somewhat problematic taxonomic past. It was treated by some as an ant of the western and eastern Mediterranean. Today it is defined as being confined to former and is not found in the latter (see the distribution and nomenclature sections for details about past treatment and the current resolution).
Identification
Güsten, Schulz and Sanetra (2006) - T. forte is most readily distinguishable from other dark, strongly sculptured Palaearctic Tetramorium species by its wide petiolar nodes. While this is not as obvious as in gynes, the postpetiole shows a conspicuously angular lateral outline in dorsal view, and the values for WI-A and WI-B are larger while that for PPL/PPW is smaller than in the most similar species, with some overlap.
Except for this character, workers of Tetramorium chefketi are very similar to T. forte, though the mesosoma is narrower and the sculpture overall more strongly rugose, particularly near the occipital corners where there is also some anastomosing of the rugae. Tetramorium moravicum workers are also similar—they may be identified by a more prominent anterio-dorsal carina at the base of the scape than in T. forte and T. chefketi, which extends into a conspicuous dorsally projecting flange. Also, in contrast to T. forte, the scape is reticulate or faintly longitudinally rugose in T. moravicum, and the occipital corners are quite prominent with the main rugae on the head running parallel throughout their length and not converging into an arcuate pattern in lateral view as in T. forte (see Schulz 1996, p. 407). Tetramorium alternans is a smaller species than T. forte with a lighter, reddish-brown colour. The scapes are shorter with a densely striate to granulate sculpture. While there are no smooth and shining spots on the waist segments, densely reticulate microsculpture predominates with only a sparse weak rugosity. In this character, T. alternans recalls Tetramorium brevicorne Bondroit, 1918 from the Tyrrhenian Islands (see Sanetra et al. 1999) rather than T. forte, T. chefketi or T. moravicum.
Workers of Tetramorium caespitum s.l. strongly differ from those discussed before by conspicuous smooth and shining medial areas on the petiolar segments (which, however, may greatly vary in width), and the head surface has a much more weakly developed rugosity, appearing shining through the lack of microsculpture. The waist segments are a lot narrower than in T. forte. The workers of Tetramorium semilaeve, and many ill-defined species similar to it, are even more weakly sculptured, yellowish to light reddish-brown and much smaller than T. forte (nest means of ML always < 0.800mm, HS < 0.730mm). Workers of Tetramorium meridionale have petiole and postpetiole at least as broad as T. forte, but in other characters generally resemble Tetramorium semilaeve.
Queens In gynes, T. forte is easily distinguished from T. chefketi, T. moravicum and Tetramorium alternans by the very broad waist segments.
Tetramorium chefketi gynes are otherwise very similar, particularly in dorsal surface sculpturing, but the mesosoma is somewhat more slender (only slightly narrower numerically) and the rugosity is more pronounced. The latter is most obvious on the head with a rugoreticulum developed near the hind margin, even extending anteriolaterally beyond the eyes (see Schulz 1996, p. 407), whereas few anastomoses between the longitudinal rugae are evident in T. forte. Tetramorium moravicum gynes differ in the structure of the scape base in a similar way as workers do; in most populations they are much larger than T. forte but microgynes are the size of large T. forte gynes. The only known gyne of T. alternans has the mesonotum more tapering anteriorly, and narrower than in T. forte and less than half of its surface (medio-posteriorly) is longitudinally rugose to striate.
The gynes of T. caespitum s.l. are much larger than those of T. forte, which is associated with a relatively smaller head and a bulging mesonotum completely concealing the pronotal corners in dorsal view, and the mesosoma is smooth and shining over two thirds of its surface or throughout. Tetramorium semilaeve gynes are also weakly sculptured (usually few shallow striae on the mesonotum), and are more lightly brownish than those of T. forte, while they are of similar size. Those of T. meridionale are even more yellowish, have a conspicuous transverse striation on the occipital margin and enlarged petiolar nodes though somewhat less than T. forte.
Even though the workers of Tetramorium maurum are very dissimilar to those of T. forte and indicate the affiliation to a different species complex, the gynes surprisingly were found to be closely similar. No morphometric characters have been detected that reliably differentiate the gynes of the two species. However, T. maurum gynes are lighter in colour (reddish-brown), the dorsal border of the petiole is not emarginated medially, the rugae on the head are less pronounced, and a larger medial unsculptured surface (> 50%) occurs on the scutellum, sometimes small unsculptured areas are also present on the petiolar nodes.
Males In males, much as in gynes, T. forte is characterized by a wider petiole and postpetiole (WI-A: 0.236–0.311) than T. chefketi and T. moravicum (nest means of WI-A always < 0.240, compare also Schulz 1996, p. 412).
In most populations, T. moravicum males are larger (ML > 2.200mm) but where microgynes occur, the size is about the same as for T. forte males. In T. forte the scutellum is clearly striate, whereas T. moravicum has a more diffuse often striolo-reticulate sculpture with sometimes a shining median part. Tetramorium alternans males are unknown.
Males of T. caespitum s.l. are much larger (nest means of ML always > 2.500mm) than those of T. forte with narrower waist segments (nest means of WI-A always < 0.240), the latter also applies to those of T. semilaeve. The male of T. meridionale has not been described.
Distribution
Güsten, Schulz and Sanetra (2006) - Lopez (1991) compiled the first comprehensive list of collecting localities of T. forte on the Iberian Peninsula (also presented as a distribution map in Lopez Gomez 1988). From these data it is evident that T. forte occurs throughout Spain up to the extreme northwest, although the species might be absent from the north coast beyond the Cantabrian Mts. There is no obvious preference for areas with a stronger Mediterranean climatic influence. In the Sierra Nevada, the species occurs at least up to 2200m. Many additional Spanish records (e.g. Tinaut 1991; De Haro & Collingwood 1991, 1992; Espadaler & Suñer 1995; Espadaler 1997b; Espadaler & Roig 2001; Reyes Lopez & Garcia 2001) confirm the ecologically generalistic occurrence of the species, which also holds true for the distribution pattern in Portugal (Paiva et al. 1990; De Haro & Collingwood 1992; Tinaut & Ruano 1994; Cammell et al. 1996; Way et al. 1997; Salgueiro 2002b, 2003; present study). Menozzi (1926) and Wheeler (1926) reported T. forte from Mallorca, but its presence on the Balearic Islands should be reconfirmed due to the commonly dubious application of the name.
Abundant samples compiled from Morocco (Cagniant 1997, collecting localities not specified) show T. forte to occur in diverse habitats from sea-level up to 2000m in the north of the country (especially in the Middle Atlas), much as in southern Spain. In the south, however, it appears much more localized at higher elevations of the High Atlas. According to Csosz (in litt.), a few samples from Algeria have been traced in collections. Three workers from Ponta Delgada (São Miguel, Azores, leg. W. M. Wheeler) in NHMB had previously been determined as T. forte, but proved to belong to T. caespitum s.l. upon investigation. The ant fauna of the Azores, largely or entirely introduced, is well known (Yarrow 1967; Heinze 1986; Salgueiro 2002a) and it seems certain that no other Tetramorium species of the caespitum-group occur. On the Canary Islands, T. forte has likewise not been recorded.
The range of T. forte extends into France along the Mediterranean coast, but except for one sample in the extreme southeast (Sommer & Cagniant 1988), no reliable records other than the original description had been published prior to this study. As Bernard´s (1967) understanding of T. forte was evidently insufficient, his locality citings from the Îles d´Hyères and the Côte d´Azur need to be re-investigated. Consequently, a gyne from Cavalaire-sur-Mer (Var) in NHMB currently represents the easternmost confirmed record. The northernmost locality in the Dept. Loire indicates an inland extension along the Rhône river for more than 200km. Only recent investigations (Schulz 1996; Schlick- Steiner et al., in press; Güsten, unpubl.) have shown that the distribution of T. forte in southern France is entwined with that of T. moravicum, which is very similar in the worker morph. Current data suggest that T. moravicum occurs at xerothermic localities with less overt Mediterranean influence compared with those of T. forte. Tetramorium moravicum is usually found above 600m where its range approaches the coast, although it may inhabit lower elevations in the Dept. Alpes-Maritimes where the ocurrence of T. forte is not confirmed. The overall distribution pattern of T. forte renders likely the postglacial recolonization into its present range from an atlanto-mediterranean refuge. Resulting contact with the ecologically similar T. moravicum progressing from a pontomediterranean refuge (Schlick-Steiner et al., in press) might have impeded further spreading of both species, but this needs additional investigation.
As stated above, records of T. forte from Corsica (Casevitz-Weulersse 1974, 1990a, 1990b) were based on a different concept of the species. Our study of comprehensive samples from Corsica and Sardinia indicated that the true T. forte is not present on the Tyrrhenian Islands.
Latitudinal Distribution Pattern
Latitudinal Range: 45.383889° to 31.205833°.
| North Temperate |
North Subtropical |
Tropical | South Subtropical |
South Temperate |
- Source: AntMaps
Distribution based on Regional Taxon Lists
Palaearctic Region: Algeria, Balearic Islands, France (type locality), Gibraltar, Iberian Peninsula, Iran, Morocco, Portugal, Spain.
Distribution based on AntMaps
Distribution based on AntWeb specimens
Check data from AntWeb
Countries Occupied
| Number of countries occupied by this species based on AntWiki Regional Taxon Lists. In general, fewer countries occupied indicates a narrower range, while more countries indicates a more widespread species. |

|
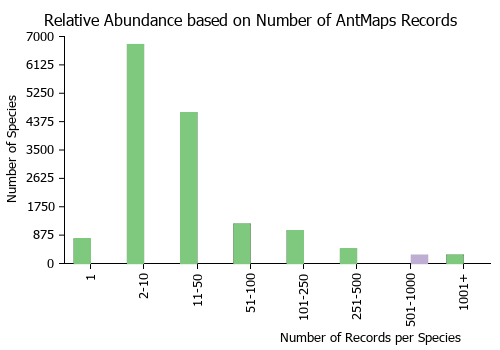
Estimated Abundance
| Relative abundance based on number of AntMaps records per species (this species within the purple bar). Fewer records (to the left) indicates a less abundant/encountered species while more records (to the right) indicates more abundant/encountered species. |
|
Biology
Güsten, Schulz and Sanetra (2006) - Apparent polygynous colonies have been observed in T. forte several times throughout its range (e.g. near Avignon, France; near Ifrane, Morocco), and the functional status as queens has been confirmed by dissection in one instance (five inseminated egg-laying queens: Sierra Nevada, Spain). Winged sexuals were recorded in T. forte colonies during late May in Spain, but during late June in the mountains of northern Portugal. The lepismatid silver-fish Proatelurina pseudolepisma (Grassi, 1887) is a generalistic myrmecophile commonly found inhabiting nests of T. forte (Molero-Baltanás et al. 1998). Astenus (Eurysunius) alcarazae Assing, 2003 and probably other species of the subgenus also occur with T. forte; these are myrmecophilous staphylinid beetles specialized to live in the colonies of ants of the genus Tetramorium in the western Palaearctic (Assing 2003). However, no records of ant social parasites collected together with T. forte are available, including the inquilines Strongylognathus testaceus and Tetramorium atratulum, which are known to use a relatively broad range of hosts in the genus Tetramorium (e.g. Sanetra et al. 1999; Sanetra & Buschinger 2000).
Association with Other Organisms
 Explore: Show all Associate data or Search these data. See also a list of all data tables or learn how data is managed.
Explore: Show all Associate data or Search these data. See also a list of all data tables or learn how data is managed.
- This ant was observed on larvae of Lampides boeticus that were feeding on Erophaca baetica in Sierra Morena, southern Spain. T. nigerrimum has also been associated with the butterflies Tomares ballus and Leptotes pirithous (Obregon et al. 2015).
- This species is associated with the aphids Aphis craccivora, Aphis rumicis, Brachycaudus cardui, Cinara pilicornis and Uroleucon sonchi (Saddiqui et al., 2019 and included references).
Castes
Worker
Images from AntWeb
   
| |
| Type of unavailable quadrinomial: Tetramorium caespitum caespitum hispanicum. Worker. Specimen code casent0904805. Photographer Z. Lieberman, uploaded by California Academy of Sciences. | Owned by MSNG, Genoa, Italy. |
Nomenclature
The following information is derived from Barry Bolton's Online Catalogue of the Ants of the World.
- forte. Tetramorium caespitum var. forte Forel, 1904b: 371 (w.q.m.) FRANCE. Subspecies of caespitum: Forel, 1905: 173; of ferox: Santschi, 1936c: 203. Raised to species: Collingwood, 1978: 71; Casevitz-Weulersse, 1990b: 420. Senior synonym of hispanicum, marocanum, ruginode, tingitanum: Güsten, Schulz & Sanetra, 2006: 5. [Name misspelled as jorte by Ruzsky, 1905b: 534.]
- ruginode. Tetramorium caespitum var. ruginode Stitz, 1917: 339 (w.) SPAIN. Santschi, 1931a: 9 (q.). Synonym of hispanicum: Collingwood, 1978: 71. [Collingwood gives hispanicum as senior name, but ruginode has priority: Bolton, 1995b: 413.] Raised to species: Bolton, 1995b: 413. Junior synonym of forte: Güsten, Schulz & Sanetra, 2006: 5.
- hispanicum. Tetramorium caespitum var. hispanicum Bondroit, 1918: 108 (q.) SPAIN. [First available use of Tetramorium caespitum subsp. caespitum var. hispanica Emery, 1909d: 701; unavailable name.] Bondroit, 1920a: 153 (w.). Subspecies of caespitum: Santschi, 1931a: 9. Raised to species: Bondroit, 1920a: 153; Collingwood & Yarrow, 1969: 73. Synonym of ruginode: Collingwood, 1978: 71. [Collingwood gives hispanicum as senior synonym, but ruginode has priority and is first available name for this taxon: Bolton, 1995b: 408.] Junior synonym of forte: Güsten, Schulz & Sanetra, 2006: 5. See also: López, 1991a: 153.
- tingitanum. Tetramorium maurum st. tingitanum Santschi, 1929e: 150 (w.q.) MOROCCO. [First available use of Tetramorium caespitum st. maura var. tingitana Santschi, 1921b: 434; unavailable name.] Junior synonym of biskrense: Cagniant, 1997: 97; of forte: Güsten, Schulz & Sanetra, 2006: 5.
- marocanum. Tetramorium marocanum de Haro & Collingwood, 1994: 101. [First available use of Tetramorium caespitum st. ferox var. marocana Santschi, 1921e: 170 (w.) MOROCCO; unavailable name. T. ferox var. marocanum Cagniant, 1964: 106; unavailable name.] Senior homonym and synonym of marocana Cagniant (below): new synonym (unpublished). Junior synonym of forte: Güsten, Schulz & Sanetra, 2006: 5.
- marocana. Tetramorium ruginode subsp. marocana Cagniant, 1997: 92 (w.q.m.) MOROCCO. Junior homonym and synonym of marocanum de Haro & Collingwood (above), hence junior synonym of forte, sensu Güsten, Schulz & Sanetra, 2006: 5.
Type Material
Güsten, Schulz and Sanetra (2006) - 1 worker, lectotype of T. caespitum forte Forel (hereby designated): T. caespitum L. worker v. forte Forel, type, Albaron Camargum / Lectotype Poldi 74 / Lectotypus Tetramorium caespitum forte Forel des. R. Güsten, A. Schulz & M. Sanetra 2005“ (Musee d'Histoire Naturelle Genève, together with 2 paralectotype workers on same pin, lectotype marked by red cardboard square); 34 paralectotype workers, Albaron (Camargue) (MHNG, 2 of these on same pin as lectotype); 2 paralectotype workers, same data as previous (Museo Civico di Storia Naturale, Genoa); 3 paralectotype workers, same data as previous (Naturhistorisches Museum, Basel); 2 paralectotype workers, same data as previous (DSTA); 11 workers, FRA, Camargue (MHNG); 3 workers, same data as previous (NHMB); 1 workers, same data as previous (DSTA); 5 workers, FRA, Albaron, 23.I.1925, leg. A. Chobaut (DSTA); 5 workers, FRA, Banyuls, leg. Saulcy (MCSN); 1 &, FRA, Var, Cavalairesur-Mer, VI.1922, leg. L. Berland (NHMB).
We have marked the lectotype by a red square with the indication “LT” attached to the cardboard triangle bearing the specimen.
Taxonomic Notes
Güsten, Schulz and Sanetra (2006):
Descriptions of T. forte gynes have been published by Bondroit (1920, as Tetramorium hispanicum), Santschi (1921b, as “T. caespitum st. maura var. tingitana”), Santschi (1932, as “T. caespitum st. hispanicum var. ruginodis”) and Cagniant (1997, as Tetramorium ruginode marocana), the latter providing a drawing of the petiolar segments. The gynes from Spain studied by Santschi (1932) are present in NHMB.
The male of T. forte had hitherto only been described by Cagniant (1997, under the name T. ruginode marocana), based on one specimen. This work included detailed drawings of genitalic characters.
The incorporation of a western Mediterranean and an eastern European taxonomic species under the nominal taxon Tetramorium forte dates back to the original description and has persisted until the present. The type series on which Forel (1904a) based his new taxon was both varied and ambiguously delimited by him. He included workers from several localities in southern France and one series of workers from the Crimean Peninsula. Gynes and males from Crimea and Transcaucasia were described; the latter were only doubtfully assigned to the new taxon. The author made conflicting statements in different sections in the description whether the sexuals from Crimea were definitely or conditionally included. Also, after describing those males and gynes, only two gynes from Crimea and one from Transcaucasia were actually listed among the specimens, but no males. Because of the reservations in assigning the sexuals to the new taxon, these are not to be considered syntypes (ICZN § 72.4.1.). According to Radchenko (1992), the sexuals in question, preserved at the Zoological Institute of the Russian Academy of Sciences, St. Petersburg, Russia (ZISP), are referable to T. caespitum (Linnaeus, 1758). The syntype series to be appraised thus consists of at least 55 workers from 4 localities in southern France (mainly at MHNG, but some also at MCSN, DSTA and ZMHB, more may be detected in other collections) along with 15 workers from Alušta, Crimea, Ukraine, of which 11 are currently deposited at ZISP (Csosz et al., unpubl.).
As a consequence of the description of the taxon from widely scattered localities preserved at different institutions, researchers in western and eastern Europe have tended to use T. forte for species from their respective areas of investigation, without addressing the inconsistency of the type series. For example, Casevitz-Weulersse (1990a) and López (1991) referred to subsets of Forel´s original specimens from southern France as types, even mentioning a putative “lectotype”, while Radchenko (1992) cited as syntypes only the specimens from Crimea and Transcaucasia, disregarding the French part of the type series. No lectotype of T. forte has ever been formally designated.
To terminate this ambiguous and instable nomenclatural situation, we decided to have the western European taxonomic species bear the name T. forte which is represented in the type series by numerous syntypes from Albaron (Camargue, France). This choice is the one most furthering stability and universality in nomenclature, following the predominant usage in the ant literature of the past 100 years including recent important phylogenetic studies. And it is also the interpretation most consistent with the original author’s intentions, because his statement of the postpetiole being about twice as wide as long only applies, in approximation, to the chosen taxonomic species among those represented in the type series.
Numerous publications have used the name T. forte in the sense coinciding with our concept (e.g. Forel 1905; Bondroit 1918; Santschi 1921a, 1921c, 1937; Bernard 1967; Collingwood & Yarrow 1969; Collingwood 1978; López 1991; Sanetra et al. 1994; Sanetra & Buschinger 2000; Steiner et al. 2005; Schlick-Steiner et al. 2005). This includes all works of the past twelve years which for the first time elucidate phylogenetic relationships in the genus Tetramorium in western and central Europe. A lesser number of publications has applied the name T. forte to eastern European and Middle Eastern species (e.g. Wheeler & Mann 1916; Agosti & Collingwood 1987a, 1987b; Radchenko 1992; Atanassov & Dlussky 1992, Arakelian 1994). However, the Tetramoriini of these parts of the Palaearctic are both very diverse and particularly little known, so that it is difficult to determine which concepts really form the basis for the mentioned usages of the name T. forte. It is likely that few, if any, refer exclusively to the taxonomic species to which the workers from Alušta (Crimea) in the syntype series of T. forte actually belong. According to current revisionary work (Csosz et al., unpubl.), T. chefketi Forel, 1911 and T. caespitum sarkissiani Forel, 1911, two names of equal priority, are available for that species, the former of which having also been used in recent taxonomic, faunistic and phylogenetic studies (Schulz 1996; Sanetra & Buschinger 2000; Schulz & Sanetra 2002; Schlick-Steiner et al. 2005; see also Appendix A). Thus it would have been a substantial disservice to nomenclatural stability to choose the lectotype of T. forte from the Alušta specimens.
A few publications, both old and recent, have used T. ruginode Stitz, 1917 as the name of the western Mediterranean species treated here (e.g. Menozzi 1926; Santschi 1932; Cagniant 1997; De Haro & Collingwood 1997; Espadaler 1997a; Salgueiro 2002a), which would have become its valid name, had T. forte been formally stabilized for the eastern European species involved in the type series. However, the name most frequently in use for the western Mediterranean species in recent publications has been T. hispanicum Bondroit, 1918 (e.g. Acosta Salmerón et al. 1983; Ortiz & Tinaut 1988; De Haro & Collingwood 1988, 1991, 1992; Paiva et al. 1990; Tinaut 1991; Espadaler & Suñer 1995; Cammell et al. 1996; Way et al. 1997; Molero-Baltanás et al. 1998; Reyes López & García 2001), and that is definitely a junior synonym of T. ruginode (see in the following section). Many of the afore-mentioned authors wrongly credited the description of T. hispanicum to Emery (1909).
The use of T. ruginode and T. hispanicum for the species probably originated in deviant concepts for T. forte which were based at least in part on type studies disregarding the Albaron specimens (Casevitz-Weulersse 1974, 1990a, 1990b; López Gómez 1988; López 1991). Our investigations revealed that at least two other taxonomic species are represented among the French syntypes. Three workers each from Nice and Palavas (near Montpellier) are relatively robust and strongly sculptured specimens of T. caespitum sensu lato (see Appendix A). They probably belong to an as yet weakly defined species separate from T. caespitum (Linnaeus, 1758) which commonly occurs along Mediterranean coasts (Schlick-Steiner et al., 2006). Study of these syntypes was probably partly responsible for the recording of T. forte from Corsica (Casevitz-Weulersse 1990a, 1990b) and for its treatment as merely a variety (Casevitz-Weulersse 1974) or subspecies (Cagniant 1997) of T. caespitum. While the postpetiole is on average slightly wider in these specimens than in true T. caespitum, it is by far not twice as wide as long as mentioned in the original description of T. forte, and rugosity and microsculpture are much less developed than in the lectotype from Albaron. Six workers from Dieulefit (Dept. Drôme) do show that pronounced sculpturing but the postpetiole is in no way shorter or wider than e.g. in T. caespitum. These workers belong to T. moravicum Kratochvil, 1941, a chiefly eastern European species described from the Czech Republic (Novak & Sadil 1941) which only recently has been found to occur in southeastern and eastern France (Schlick-Steiner et al., in press; Güsten, unpubl.). The addition of these specimens to the syntype series by Forel (1904a) can be ascribed to the incomplete understanding of variability and critical characters in the genus at the time. Unless otherwise noted the text for the remainder of this section is reported from the publication that includes the original description.
Description
Güsten, Schulz and Sanetra (2006):
Worker
Measurements and indices (n=34): HL 0.824±0.057(0.725–0.936)mm, HW 0.783±0.059(0.680–0.906)mm, HS 0.804±0.056(0.702–0.921)mm, SL 0.631±0.038(0.563–0.728)mm, ML 0.986±0.111(0.831–1.194)mm, MW 0.529±0.046(0.456–0.637)mm, PSL 0.104±0.016(0.076–0.143)mm, PEL 0.328±0.033(0.247–0.385)mm, PEW 0.290±0.030(0.219–0.342)mm, PEH 0.271±0.026(0.238–0.323)mm, PPL 0.208±0.017(0.171–0.238)mm, PPW 0.335±0.037(0.257–0.404)mm, HW/HL 0.951±0.022(0.912–1.020), SL/HS 0.786±0.026(0.727–0.840), MW/ML 0.560±0.032(0.509–0.675), PSL/ML 0.110±0.012(0.085–0.130), PEH/PEL 0.826±0.048(0.750–1.019), PEW/PEL 0.868±0.078(0.742–1.192), PEW/HS 0.360±0.019(0.311–0.402), PPL/PPW 0.622±0.044(0.553–0.688), PPW/HS 0.419±0.021(0.365–0.466), PEW/PPW 0.860±0.044(0.813–1.033), WI-A 0.322±0.019(0.269–0.359), WI-B 0.390±0.018(0.338–0.430).
Measurements and indices of the lectotype: HL 0.906mm, HW 0.891mm, HS 0.898mm, SL 0.675mm, ML 1.102mm, MW 0.618mm, PSL 0.143mm, PEL 0.385mm, PEW 0.328mm, PEH 0.323 mm, PPL 0.238 mm, PPW 0.394mm.
Larger Palaearctic Tetramorium worker with subquadrate head. Preoccipital margin nearly straight to concave, genae more or less straight, outlines convergent. Head widest behind the eyes. Mesosoma robust, broad, with pronounced pronotal angles. Mesopropodeal suture shallowly depressed. Propodeal spines moderately long and straight. Petiole robust, node in lateral view rather rounded, outline anterior of node concave. Petiole and postpetiole broad in relation to mesosoma, postpetiole with laterally prominently protruding angles. Dark brown to blackish, appendages lighter, orange-brown. Head, dorsal parts of mesosoma, petiole and postpetiole entirely carinate or rugose. Frontal area of head with 14–16 even rugae which diverge slightly towards the preoccipital margin, converging into a conspicuously arcuate pattern in lateral view (see Schulz 1996, p. 407). Genae and surface of occipital corners rugose. Dorsal surface of head with reticulate microsculpture, but with few more conspicuous anastomoses between principal rugae. Ventral head surface longitudinally striate without any microsculpture. Scapes usually smooth and shinning, sometimes with diffuse microsculpture, and with an inconspicuous anterio-dorsal carina at the base which may grade into the trace of a transverse extension but not into a conspicuous dorsally projecting flange. Dorsal surface of mesosoma rugose with variably developed reticulate microsculpture, on the propodeum evenly and roughly reticulate, especially between the spines. Dorsal part of petiole and postpetiole longitudinally to concentrically, often rather irregularly rugose with reticulate microsculpture, no weakening of sculpture on dorsalmost surfaces. Ventral parts of petiolar nodes heavily reticulate. Polygonal microsculpture on the first gaster tergite never absent, rarely covers the whole surface of the tergite (in some Moroccan specimens). On the anteriormost part of the tergite, this microsculpture can appear striated in some specimens. Frequency of the latter feature within the same nest series increases towards the south of the species’ range.
Queen
Measurements and indices (n=23): HL 1.064±0.073(0.842–1.293)mm, HW 1.127± 0.096(0.891–1.391)mm, HS 1.096±0.080(0.866–1.330)mm, SL 0.791±0.044(0.634–0.861)mm, ML 1.762±0.101(1.391–1.879)mm, MW 1.082±0.074(0.830–1.196)mm, PSL 0.147±0.022(0.105–0.181)mm, PEL 0.467±0.029(0.380–0.504)mm, PEW 0.559±0.047(0.418–0.618)mm, PEH 0.460±0.034(0.371–0.518)mm, PPW 0.711±0.054(0.556–0.817)mm, HW/HL 1.060±0.060(1.000–1.326), SL/HS 0.723±0.034(0.602–0.763), HS/ML 0.622±0.028(0.581–0.727), MW/ML 0.614±0.019(0.568–0.653), PSL/ML 0.083±0.011(0.062–0.102), PEH/PEL 0.984±0.059(0.902–1.111), PEW/PEL 1.194±0.097(1.000–1.383), PEW/HS 0.511±0.037(0.421–0.569), PPW/HS 0.650±0.045(0.525–0.712), PEW/PPW 0.787±0.042(0.630–0.855), WI-A 0.360±0.017(0.324–0.392).
Medium-sized Palaearctic Tetramorium gyne, generally with rather robust appearance. Head with rather rounded preoccipital corners and straight to slightly convex, somewhat convergent genal outlines. Scape relatively short and broad. Mesosoma short and robust, with flat (not bulging) dorsal outline. In dorsal view the pronotal angles are fully visible. Propodeal spines broadly attached, triangular with pointed tips, orientation subcaudate. Petiole and postpetiole very wide, lobe-like, the petiole medially emarginated. First gaster tergite with at least a few erect hairs. Colour as in workers. Frons rugose, the rugae divergent and curving towards the occipital corners with little or no anastomosing. Genae rugose, ventral head surface longitudinally striate. On the genae and near the occipital corners, a fine reticulate microsculpture occurs between the main rugae. Sides of mesosoma and petiolar segments mainly longitudinally carinate, restricted parts only rugose. In dorsal view, pronotum with rugose sculpture, mesonotum longitudinally rugose but more weakly so laterally, with a very small smooth and shining spot anterio-medially, scutellum rugose except for narrow smooth median part. Sculpturing between the spines variable, principally longitudinally rugose. Sculpture of dorsal surface of waist segments also variable, diffusely rugose to rugulose, to concentrically striate. Individuals with more pronounced sculpturing have the rugose portion more strongly developed. Polygonal microsculpture covers small spots on the first gaster tergite, appearing longitudinally striate on the anterior part (0.150–0.250mm) of the tergite.
Male
Measurements and indices (n=22): HL 0.738±0.019(0.702–0.770)mm, HW 0.752±0.043(0.687–0.891)mm, HS 0.745±0.025(0.695–0.800)mm, SL 0.344±0.011(0.323–0.361)mm, 2FL 0.406±0.018(0.361–0.428)mm, ED 0.278±0.012(0.257–0.304)mm, ML 2.028±0.064(1.891–2.135)mm, MW 1.210±0.064(1.098–1.318)mm, PEW 0.482±0.048(0.390–0.589)mm, PPW 0.637±0.046(0.570–0.722)mm, HW/HL 1.020±0.059(0.959–1.255), SL/HS 0.462±0.022(0.421–0.501), SL/2FL 0.847±0.041(0.778–0.925), MW/ML 0.597±0.037(0.538–0.663), PEW/HS 0.647±0.057(0.529–0.772), PPW/HS 0.854±0.051(0.778–0.934), PEW/PPW 0.758±0.056(0.661–0.848), WI-A 0.276±0.017(0.236–0.311).
Small Palaearctic Tetramorium male, with broad head and relatively large eyes. Mesonotum and scutellum bulging. Propodeal spines well visible, but short and more or less triangular, tooth-like. Petiole and postpetiole very broad, petiole on each side with two laterally oriented processes and a distinctly emarginate median part. Isolated erect hairs on first gaster tergite. Colour dark brown, appendages yellowish orange. Sculpture on head, mesosoma and waist dense. Head largely rugoreticulate, pronotum and lateral parts of mesosoma chiefly longitudinally striate with reticulate microsculpture, mesonotum longitudinally to concentrically striate but with extensive parts laterally and anterio-medially smooth and shining. Scutellum completely striate, propodeum diffusely striate to reticulate, waist segments reticulate, gaster without sculpture.
Karyotype
- See additional details at the Ant Chromosome Database.
- Explore: Show all Karyotype data or Search these data. See also a list of all data tables or learn how data is managed.
- n = 14, karyotype = 4SM+3ST+7A (Spain) (Palomeque et al., 1987; Lorite et al., 2000) (as T. hispanicum).
References
- Gusten, R., A. Schulz and M. Sanetra. 2006. Redescription of Tetramorium forte Forel, 1904 (Insecta: Hymenoptera: Formicidae), a western Mediterranean ant species. Zootaxa. 1310:1-35.
- Arcos, J., Chaves, D., Alarcón, P., Rosado, A. 2022. First record of Temnothorax convexus (Forel, 1894) in Portugal (Hymenoptera: Formicidae) with an updated checklist of the ants from the country. Sociobiology, 69(2), e7623 (doi:10.13102/sociobiology.v69i2.7623).
- Arcos, J., Chaves, D., Alarcón, P., Rosado, Á. 2022. First record of Temnothorax convexus (Forel, 1894) in Portugal (Hymenoptera: Formicidae) with an updated checklist of the ants from the country. Sociobiology, 692), e7623 (doi:10.13102/sociobiology.v69i2.7623).
- Atanassov, N.; Dlussky, G. M. 1992. Fauna of Bulgaria. Hymenoptera, Formicidae. Fauna Bûlg. 22: 1-310 (page 152, Senior synonym of moravicum and taurocaucasicum)
- Bernard, F. 1967a [1968]. Faune de l'Europe et du Bassin Méditerranéen. 3. Les fourmis (Hymenoptera Formicidae) d'Europe occidentale et septentrionale. Paris: Masson, 411 pp. (page 233, Senior synonym of moravicum)
- Borowiec, L. 2014. Catalogue of ants of Europe, the Mediterranean Basin and adjacent regions (Hymenoptera: Formicidae). Genus (Wroclaw) 25(1-2): 1-340.
- Bracko, G., Wagner, H.C., Schulz, A., Gioahin, E., Maticic, J., Trantnik, A. 2014. New investigation and a revised checklist of the ants (Hymenoptera: Formicidae) of the Republic of Macedonia. North-Western Journal of Zoology 10: 10-24.
- Casevitz-Weulersse, J. 1990c. Étude systématique de la myrmécofaune corse (Hymenoptera, Formicidae) (Deuxième partie). Bull. Mus. Natl. Hist. Nat. Sect. A Zool. Biol. Écol. Anim. (4) 12: 415-442 (page 420, Raised to species)
- Catarineu, C., Barberá, G.G., Reyes-López, J.L. 2018. Zoogeography of the ants (Hymenoptera: Formicidae) of southeastern Iberian Peninsula. Sociobiology 65, 383-396 (doi:10.13102/sociobiology.v65i3.2822).
- Collingwood, C. A. 1978. A provisional list of Iberian Formicidae with a key to the worker caste (Hym. Aculeata). EOS. Rev. Esp. Entomol. 52: 65-95 (page 71, Raised to species, Senior synonym of silvestrianum)
- Dlussky, G. M.; Soyunov, O. S.; Zabelin, S. I. 1990 [1989]. Ants of Turkmenistan. Ashkhabad: Ylym Press, 273 pp. (page 202, Senior synonym of moravicum and taurocaucasicum)
- Forel, A. 1904c [1903]. Note sur les fourmis du Musée Zoologique de l'Académie Impériale des Sciences à St. Pétersbourg. Ezheg. Zool. Muz. 8: 368-388 (page 371, worker, queen, male described)
- Forel, A. 1905a. Einige biologische Beobachtungen des Herrn Prof. Dr. E. Göldi an brasilianischen Ameisen. Biol. Centralbl. 25: 170-181 (page 173, Race/stirps of caespitum)
- Obregon, R., M. R. Shaw, J. Fernandez-Haeger, and D. Jordano. 2015. Parasitoid and ant interactions of some Iberian butterflies (Insecta: Lepidoptera). Shilap-Revista De Lepidopterologia. 43:439-454.
- Radchenko, A. G. 1992b. Ants of the genus Tetramorium (Hymenoptera, Formicidae) of the USSR fauna. Report 2. Zool. Zh. 71(8 8: 50-58 (page 51, Senior synonym of moravicum and taurocaucasicum)
- Salata, S., Khalili-Moghadam, A., Borowiec, L. 2024. A new species of the Tetramorium meridionale species-group (Hymenoptera: Formicidae) from Iran. Zoology in the Middle East 70, 161–175 (doi:10.1080/09397140.2024.2359167).
- Sanetra, M., Buschinger, A. 2000. Phylogenetic relationships among social parasites and their hosts in the ant tribe Tetramoriini (Hymenoptera: Formicidae). European Journal of Entomology 97: 95-117.
- Santschi, F. 1936c. Liste et descriptions de fourmis du Maroc. Bull. Soc. Sci. Nat. Maroc 16: 198-210 (page 203, Race/stirps of ferox)
- Siddiqui, J. A., Li, J., Zou, X., Bodlah, I., Huang, X. 2019. Meta-analysis of the global diversity and spatial patterns of aphid-ant mutualistic relationships. Applied Ecology and Environmental Research 17: 5471-5524 (doi:10.15666/aeer/1703_54715524).
References based on Global Ant Biodiversity Informatics
- Abril S., and C. Gomez. 2013. Rapid assessment of ant assemblages in public pine forests of the central Iberian Peninsula. Forest Ecology and Management 293: 7984.
- Acosta, F. J., M. D. Martínez Ibáñez, and M. A. Morales. "Contribución al conocimiento de la mirmecofauna del encinar peninsular. I." Boletín de la Asociación Española de Entomología 6 (2) (1983): 379-391.
- Agosti, D. and C.A. Collingwood. 1987. A provisional list of the Balkan ants (Hym. Formicidae) and a key to the worker caste. I. Synonymic list. Mitteilungen der Schweizerischen Entomologischen Gesellschaft, 60: 51-62
- Aldawood AS, Sharaf MR (2011) Monomorium dryhimi sp. n., a new ant species (Hymenoptera, Formicidae) of the M. monomorium group from Saudi Arabia, with a key to the Arabian Monomorium monomorium-group. ZooKeys 106: 4754. doi: 10.3897/zookeys.106.139
- AntArea. Accessed on February 5th 2014 at http://antarea.fr/fourmi/
- Antarea (at www.antarea.fr on June 11th 2017)
- Asociacion Iberica de Mirmecologia. 2011. List of species collected during the Taxomara Lisboa 2010. Iberomyrmex 3: 32-33.
- Asociacion Iberica de Mirmecologia. 2014. List of species collected during the Taxomara 2014 Oviedo. Iberomyrmex 6: 23-24.
- Azcarate F. M., and B. Peco. 2012. Abandonment of grazing in a mediterranean grassland area: consequences for ant assemblages. Insect Conservation and Diversity 5: 279288.
- Bernard F. 1967. Faune de l'Europe et du Bassin Méditerranéen. 3. Les fourmis (Hymenoptera Formicidae) d'Europe occidentale et septentrionale. Paris: Masson, 411 pp.
- Berville L., C. Santelli, J. Reybaud, M. Renucci, P. Ponel, O. Blight, and E. Provost. 2014. Suivi d’un site atelier dans le golfe de Fos: Une diversite myrmecologique insoupconne. Etudes Vauclusiennes 82: 71-78.
- Bondroit, J. "Notes diverses sur des fourmis d'Europe." Annales de la Société Entomologique de Belgique 59 (1920): 143-158.
- Borowiec L. 2014. Catalogue of ants of Europe, the Mediterranean Basin and adjacent regions (Hymenoptera: Formicidae). Genus (Wroclaw) 25(1-2): 1-340.
- Borowiec L., and S. Salata. 2017. New records of ants (Hymenoptera: Formicidae) from southern Portugal. Acta Entomologica Silesiana 25(3): 1-10.
- Cagniant H. 1997. Le genre Tetramorium au Maroc (Hymenoptera: Formicidae): clé et catalogue des espèces. Ann. Soc. Entomol. Fr. (N.S.) 33: 89-100.
- Cagniant, H. 2006. Liste actualisée des fourmis du Maroc. Myrmecologische Nachrichten 8: 193-200.
- Cagniant, H., and X. Espadaler. "Liste des espèces de fourmis du Maroc." Actes des Colloques Insectes Sociaux 8 (1993): 89-93.
- Cagniant, H. "Contribution à la connaissance des fourmis marocaines. Aphaenogaster espadaleri n. sp. (Hymenoptera, Myrmicidae)." Nouvelle Revue d'Entomologie (n.s.)1 (1984): 387-395.
- Cagniant, H. "Contribution à la connaissance des fourmis marocaines. Description des trois castes d'Aphaenogaster wilsoni n. sp. (Hymenoptera, Myrmicidae)." Nouvelle Revue d'Entomologie (n.s.)5 (1988): 49-55.
- Cagniant, H. "Contribution à la connaissance des fourmis marocaines: description des sexués et compléments à la définition de l'espèce Aphaenogaster theryi Santschi 1923 (Hyménoptères, Formicoidea, Myrmicidae)." Bulletin de la Société d'Histoire Naturelle de Toulouse 122 (1986): 139-144.
- Cagniant, H. "Liste actualisee des fourmis du Maroc (Hymenoptera: Formicidae)." Myrmecologische Nachrichten 8 (2006): 193-200.
- Carpintero S., J. Reyes-Lopes, and G. M. Luque. 2011. Ant community structure under Retama sphaerocarpa shrubs in a semi-arid environment. Entomological Science 14: 147153.
- Carpintero, S., A. Tinaut, J. Reyes, and L. Arias. "Estudio faunístico de los formícidos (Hymenoptera: Formicidae) del Parque Nacional de Doñana." Boletín de la Asociación Española de Entomología 25 (1-2) (2001): 133-152.
- Casevitz-Weulersse J. 1990. Etude Systematique de la Myrmecofaune Corse (Hymenoptera: Formicidae), deuxieme partie. Bull. Mus. Natn. Hist. Nat. Paris. 4eme serie 12, section A(2): 415-442.
- Casevitz-Weulersse J., and C. Galkowski. 2009. Liste actualisee des Fourmis de France (Hymenoptera, Formicidae). Bull. Soc. Entomol. Fr. 114: 475-510.
- Casevitz-Weulersse, J. "Contribution a la connaisance des fourmis de la Corse (Hymenoptera: Formicidae)." These de Doctorat Museum Nat (1989): 379pp.
- Collingwood C., and A. Prince. 1998. A guide to ants of continental Portugal (Hymenoptera: Formicidae). Boletim da Sociedade Portuguesa de Entomologia. Suplemento 5: 1-49.
- Collingwood, C. A., and I. H. H. Yarrow. "A survey of Iberian Formicidae." EOS (Revista española de entomología) 44 (1969): 53-101.
- Collingwood, C. A. "A provisional list of Iberian Formicidae with a key to the worker caste." EOS (Revista española de entomología) Nº LVII (1978): 65-95.
- Comín del Río, P. Estudio de los formícidos de Baleares: Contribución al estudio taxonómico, geográfìco y biológico. Palma de Mallorca: Universidad de las Islas Baleares, 1988.
- Delye G. 1983. Contribution a l'etude des peuplements des invertebres de milieux extremes: les fourmis des dunes littorales del aCamargue (B. d. Rh) et de l'Espiguette (Gard). Bull. Soc. Linneenne Provence 35: 121-124.
- Diniz, M. A. "Estado actual do conhecimento dos himenópteros de Portugal." Memorias e Estudos do Museu Zoologico da Universidade de Coimbra 259 (1959): 1-42.
- Dlussky G. M., O. S. Soyunov, and S. I. Zabelin. 1990. Ants of Turkmenistan. Ashkabad: Ylym Press, 273 pp.
- Eidmann, H. "Die Ameisenfauna der Balearen." Zeitschrift für Morphologie und Ökologie der Tiere 6 (1926): 694-742.
- Eidmann, H. "Zur Kenntnis der Insektekfauna der Balearischen Inseln." Entomologische Mitteilungen 16 (1927): 24-37.
- Emery, C. "Beiträge zur Monographie der Formiciden des paläarktischen Faunengebietes. (Hym.) Teil IX. Tetramorium." Deutsche Entomologische Zeitschrift 1909 (1909): 695-712.
- Emery, C. "Hymenoptera. Fam. Formicidae. Subfam. Myrmicinae. [part]." en Wytsman, P. "Genera Insectorum" 174B (1922): 95-206.
- Espadaler X. F. Garcia, K. Gomez, S. Serrano, and R. Vila. 2009. Ants (Hymenoptera, Formicidae) from Mont-Rebei gorge (Pallars Jussà). Boletín Sociedad Entomológica Aragonesa 44: 393399.
- Espadaler X., and J. I. López-Colón. 2011. Hormigas (Hymenoptera, Formicidae) de una zona de yesos de la Comunidad de Madrid (España) Boletín de la Sociedad Entomológica Aragonesa 49: 261-264.
- Espadaler, X. "Catalogos entomofauna aragonesa: Familia: Formicidae." Catalogos entomofauna aragonesa 13 (1997): 13-21.
- Espadaler, X. "Formigues del garraf: Coneixement actual i clau d'identificació." I trobada d'estudiosos de garraf: Monografies 19 (1992): 9-13.
- Espadaler, X. "Formícidos de los alrededores de la laguna de Sariñena (Huesca). Descripción del macho de Camponotus foreli Emery." Colección Estudios Altoaragoneses 6 (1986): 109-126.
- Espadaler, X. "Lasius neglectus Van Loon, Boosma & Andrásfalvy, 1990 a potential pest ant in Spain." Orsis 14 (1999): 43-46.
- Forel A. 1904. Note sur les fourmis du Musée Zoologique de l'Académie Impériale des Sciences à St. Pétersbourg. Ezheg. Zool. Muz. 8: 368-388.
- Forel A. 1905. Miscellanea myrmécologiques II (1905). Ann. Soc. Entomol. Belg. 49: 155-185.
- Forel, A. "Miscellanea myrmécologiques II (1905)." Annales de la Société Entomologique de Belgique 49 (1905): 155-185.
- GRETIA. 2017. Bilan annuel de l'enquete sur la repartition des fourmis armoricaines. 23 pages.
- Garcia Garcia F., and A. D. Cuesta-Esgura. 2017. First catalogue of the ants of Burgos province, Spain (Hymenoptera: Formicidae). Boletín de la Sociedad Entomológica Aragonesa 60: 245–258.
- Gomez C., D. Casellas, J. Oliveras, and J. M. Bas. 2003. Structure of ground-foraging ant assemblages in relation to land-use change in the northwestern Mediterranean region. Biodiversity and Conservation 12: 21352146.
- Gomez K., D. Martinez, and X. Espadaler. 2018. Phylogeny of the ant genus Aphaenogaster (Hymenoptera: Formicidae) in the Iberian Peninsula, with the description of a new species. Sociobiology 65: 215-224.
- Gouraud C. 2015. Bilan de l’année 2014 : Atlas des fourmis de Loire-Atlantique (Hymenoptera, Formicidae). Atlas des Formicidae de Loire-Atlantique, compte rendu de la première année d’étude (2014)
- Gratiashvili N., Barjadze S. 2008. Checklist of the ants (Formicidae Latreille, 1809) of Georgia. Proceedings of the Institute of Zoology (Tbilisi) 23: 130-146.
- Gómez C., and S. Abril. 2011. Selective logging in public pine forests of the central Iberian Peninsula: Effects of the recovery process on ant assemblages Forest Ecology and Management 262: 1061-1066.
- Güsten, R., A. Schulz, and M. Sanetra. "Redescription of Tetramorium forte Forel, 1904 (Insecta: Hymenoptera: Formicidae), a western Mediterranean ant species." Zootaxa 1310 (2006): 1-35.
- Karaman M. G. 2009. An introduction to the ant fauna of Macedonia (Balkan Peninsula), a check list (Hymenoptera, Formicidae). Natura Montenegrina 8(3): 151-162.
- Karaman M. G. 2011. A catalogue of the ants (Hymenoptera, Formicidae) of Montenegro. Podgorica: Catalogues 3, Volume 2, Montenegrin Academy of Sciences and Arts, 140 pp.
- Karaman, M. G. "Zoogeography, diversity and altitudinal distribution of ants (Hymenoptera: Formicidae) in the Mediterranean and the oro-Mediterranean parts of Montenegro." NORTH-WESTERN JOURNAL OF ZOOLOGY 7, no. 1 Article No.: 101105 (2010): 1-9.
- Kiran K., and N. Aktac. 2006. The vertical distribution of the ant fauna (Hymenoptera: Formicidae) of the Samanh Mountains, Turkey. Linzer Biol. Beitr. 38(2): 1105-1122.
- Legakis Collection Database
- Lorite P., J. A. Carrillo, M. F. García, and T. Palomeque. 2000. Chromosome numbers in Spanish Formicidae. III. Subfamily Myrmicinae (Hymenoptera). Sociobiology 36: 555-570.
- Luque-García, G., J. L. Reyes-López, and J. Fernández-Haeger. "Estudio faunístico de las hormigas (Hymenoptera: Formicidae) de la cuenca del río Guadiamar: Primeras aportaciones." Boletín de la Sociedad Entomológica Aragonesa 30 (2002): 153-159.
- López, F. "Estudio morfológico y taxonómico de los grupos de especies ibéricas del género Tetramorium Mayr, 1855." Boletín de la Asociación Española de Entomología 15 (1991): 29-52.
- Markó B. 1997. Contribution to the Knowledge of the Ant-Fauna (Hymenoptera: Fromicoidea) of the Cri?ul-Repede River-Valley. In: Sárkány-Kiss, A., Hamar, J. (szerk.): The Cri?/Körös Rivers' Valleys. A study of the geography, hydrobiology and ecology of the river system and its environment. Tiscia Monograph Series 2, Szolnok-Szeged-Tîrgu Mure?, pp. 345-352.
- Martínez Ibáñez, M. D., and X. Espadaler. "Revisión de las hormigas ibéricas de la colección M. Medina y nuevos datos de distribución." Actas de las VIII Jornadas de la Asociación Española de Entomología (1986): 1022-1034.
- Menozzi, C. "Zur Kenntnis der Ameisenfauna der Balearen." Zoologischer Anzeiger 66 (1926): 180-182.
- Morillo C. 1981. Inventario y fenologia de los Formicidae de un encinar de Espana central. Rev. Ecol. Biol. Sol. 18(2): 243-251.
- Notario, A., L. Castresana, and L. M. López-Arce. "Contribución al estudio de la entomofauna del monte del estado Lugar Nuevo (Jaén)." Ecología Nº 9 (1995): 447-463.
- Núñez-Pérez, E. "Bases para el desarrollo del control integrado de los pulgones (Hom. Aphididae) de los cultivos de la provincia de León." Universidad de León, Secretariado de Publicaciones Tesis doct (1992): 7 páginas y 4 microfichas. León.
- Obregon R., J. Lopez, J. L. Reyes-Lopez. 2014. A catalogue of the ant fauna (Hymenoptera, Formicidae) of Sierra Mágina (Jaén, southern Spain). Boletín de la Sociedad Entomológica Aragonesa (S.E.A.), 54: 370374.
- Ordóñez-Urbano C., J. Reyes-López, and S. Carpintero-Ortega. 2007. Estudio faunísidos de los formícidos (Hymenoptera: Formicidae) asociados a los bosques de ribera en la Provincia de Córdoba (España). Primeras Aportaciones. Boletin Sociedad Entomológica Aragonesa 40: 367-375.
- Ortiz, F. J., and A. Tinaut. "Introducción al conocimiento de las hormigas de la provincia de Almería." Publicaciones del Instituto de Estudios Almerienses. Boletín (Ciencias) 8 (1988): 223-231.
- Paraschivescu D. 1978. Elemente balcanice in mirmecofauna R. S. Romania. Nymphaea 6: 463- 474.
- Passera L. 1977. Peuplement myrmecologique du cordon littoral du Languedoc-Roussillon modifications anthropiques. Vie et Milieu 27: 249-265.
- Petrov I. Z. 2002. Contribution to the myrmecofauna (Formicidae, Hymenoptera) of the Banat Province (Serbia). Archives of Biological Sciences, Belgrade, 54(12): 57-64.
- Petrov I. Z. 2004. A list of currently known ant species (Formicidae, Hymenoptera) of Serbia. Arch. Biol. Sci., Belgrade 56(3-4): 121-125.
- Petrov I. Z. 2008. Notes on the currently known ant species (Hymenoptera: Formicidae) of Montenegro. Bulletin of the Natural History Museum 1: 243-254.
- Petrov I. Z., and C. A. Collingwood. 1992. Survey of the myrmecofauna (Formicidae, Hymenoptera) of Yugoslavia. Archives of Biological Sciences (Belgrade) 44: 79-91.
- Restrepo, C., X. Espadaler, and Andrés de Haro. "Contribución al conocimiento faunístico de los formícidos del Macizo de Garraf (Barcelona)." Orsis (Organismes i Sistemes) 1 (1986): 113-129.
- Retana J., and X. Cerdá X. 2000. Patterns of diversity and composition of Mediterranean ground ant communites tracking spatial and temporal variability in the thermal environment. Oecologia 123: 436-444.
- Reyes, J. L., and G. Luque. "Nuevos datos sobre el género Cardiocondyla (Hym.: Formicidae) en andalucía (Sur de la Península Ibérica)." Boletín de la Asociación Española de Entomología 25 (3-4) (2001): 125-126.
- Rodríguez, A. "Contribución al conocimiento de las hormigas (Hymenoptera, Formicidae) de Sierra Morena Central." Boletín de la Asociación Española de Entomología 5 (1982): 181-188.
- Roig X., X. Espadaler, R. Cusco, F. Garcia, K. Gomez, S. Serrano, and R. Vila. 2008. Ants in gypsum areas. First record from Catalonia (Iberian Peninsula) of Cardiocondyla batesii Forel (Hymenoptera, Formicidae) with an updated list of Catalan Formicidae. Boletin Sociedad Entomologica Aragonesa 42: 189-192.
- Salgueiro, J. "Primeiros registros de Formicídeos (Hymenoptera) para dúas áreas protegidas portuguesas. Adiçao de duas novas especies á mirmecofauna de Portugal." Boletín de la Sociedad Entomológica Aragonesa 32 (2003): 109-110.
- Sanchez-Gil Jimeno R., and J. L. Reyes-Lopez. 2016. Study of ants species of the Sierra de San Carlos del Valle (Ciudad Real) and updating the provincial check list (Hymenoptera, Formicidae). Boln. Asoc. esp. Ent. 40 (1-2): 93-109.
- Santschi F. 1910. Contributions à la faune entomologique de la Roumanie. Formicides capturées par Mr. A. L. Montandon. Buletinul Societatii Române de Stiinte 19: 648-652.
- Santschi, F. "Fourmis du Maroc, d'Algérie et de Tunisie." Bulletin et Annales de la Société Entomologique de Belgique (Bruxelles) 69 (1929): 138-165.
- Schlick-Steiner, B.C., F.M. Steiner, M. Sanetra, G. Heller, C. Stauffer, E. Christian and B. Seifert. 2005. Queen size dimorphism in the ant Tetramorium moravicum (Hymenoptera, Formicidae): Morphometric, molecular genetic and experimental evidence. Insects Sociaux 52:186-193
- Schmitz H. 1950. Formicidae quaedam a cl. A. Stärcke determinatae, quas in Lusitania collegit. Brotéria Ser. Cienc. Nat. 19: 12-16.
- Steiner F. M., B. Seifert, K. Moder, and B. C. Schlick-Steiner. 2010. A multisource solution for a complex problem in biodiversity research: description of the cryptic ant species Tetramorium alpestre sp. n. (Hymenoptera: Formicidae). Zoologischer Anzeiger 249: 223-254.
- Stitz, H. "Ameisen aus dem westlichen Mittelmeergebiet und von den Kanarischen Inseln." Mitteilungen aus den Zoologischen Museum in Berlin 8 (1917): 333-353.
- Suñer i Escriche, David. "Contribució al coneixement mirmecologic de Gavarres, Montgrí, Guilleríes i la Serralada Transversal." Tesis Doctoral Universida (1991): 577 pp.
- Sánchez, I., and X. Espadaler. "Una nueva población de Cataglyphis floricola Tinaut, 1993 (Hymenoptera, Formicidae)." Boletín de la Sociedad Entomológica Aragonesa 39 (2006): 318.
- Tinaut A. 2016. Ants of the Tejeda, Almijara and Alhama Mountains Natural Park (Andalusia, Spain) (Hymenoptera, Formicidae). Boln. Asoc. esp. Ent., 40 (1-2): 125-159.
- Tinaut A., and F. J. Ortiz. 1988. Introduccion al conocimiento de las hormigas de la provincia de Almeria (Hymenoptera: Formicidae). Publicaciones del Instituto de Estudios Almerienses. Boletín (Ciencias) 8: 223-231.
- Tinaut, A. "Contribución al conocimiento de los formícidos del Parque Nacional de Doñana." Boletín de la Asociación Española de Entomología 15 (1991): 57-63.
- Tizado Morales, E. J. Estudio comparado de la fauna y la biología de pulgones (Hom.), afidíinos (Hym.) y otros insectos acompañantes en dos áreas de la provincia de León In Secretariado de Publicaciones, Tesis doctoral en microficha, nº 67. León: Universidad de León, 1991.
- Tohme G. 1996. Formicidae. Etude de la diversité biologique n° 4 . Ministère de lAgriculture à Beyrouth (Eds.). P85-87.
- Tohme G., and H. Tohme. 2014. Nouvelles liste des especes de fourmis du Liban (Hymenoptera, Formicoidea). Lebanese Science Journal 15(1): 133-141.
- Trigos Peral G., and J. Reyes Lopez. 2016. Ants as bioindicators for monitoring of urban green in the south of the Iberian Peninsula. Proposal of functional groups. Iberomyrmex 8: 37-38.
- Trigos-Peral G., and J. L. Reyes-Lopez. 2013. Primera relacion de los formicidos (Hymenoptera, Formicidae) de la Reserva Natural Lagunas de Campillos (Malaga). Boln. Asoc. esp. Ent. 37 (3-4): 217-224.
- Wheeler W. M. and W. M. Mann. 1916. The ants of the Phillips Expedition to Palestine during 1914. Bulletin of the Museum of Comparative Zoology 60: 167-174.
- Wheeler W. M., and W. M. Mann. 1916. The ants of the Phillips Expedition to Palestine during 1914. Bulletin of the Museum of Comparative Zoology 60: 167-174.
- Wheeler, W. M. "Ants of the Balearic Islands." Folia Myrmecologica et Termitologica 1 (1926): 1-6.
- de Haro, Andrés, and C. A. Collingwood. "Prospección mirmecológica por Andalucia." Boletín de la Estación Central de Ecología 6 (12) (1977): 85-90.
- de Haro, Andrés, and C. A. Collingwood. "Prospección mirmecológica por Extremadura (España) y Sao Brás-Almodovar, Alcácer do Sal, Serra da Estrela (Portugal)." Boletim da Sociedade Portuguesa de Entomologia Suplemento 3(1) (1992): 95-104.
- de Haro, Andrés, and C. A. Collingwood. "Prospección mirmecológica por el litoral mediterráneo de Marruecos (Cabo Negro, Martil, Oued Lau) y su comparación con la zona meridional ibérica." Orsis 9 (1994): 97-104.
- de Haro, Andrés, and C. A. Collingwood. "Prospección mirmecológica por la península Tingitana al norte del Rif (Marruecos)." Orsis (Organismes i Sistemes) 12 (1997): 93-99.
- Pages using DynamicPageList3 parser function
- Need species key
- North temperate
- North subtropical
- Butterfly Associate
- Host of Lampides boeticus
- Host of Tomares ballus
- Host of Leptotes pirithous
- Aphid Associate
- Host of Aphis craccivora
- Host of Aphis rumicis
- Host of Brachycaudus cardui
- Host of Cinara pilicornis
- Host of Uroleucon sonchi
- Karyotype
- Species
- Extant species
- Formicidae
- Myrmicinae
- Crematogastrini
- Tetramorium
- Tetramorium forte
- Myrmicinae species
- Crematogastrini species
- Tetramorium species
- Ssr