Pogonomyrmex huachucanus
| Pogonomyrmex huachucanus | |
|---|---|

| |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Arthropoda |
| Class: | Insecta |
| Order: | Hymenoptera |
| Family: | Formicidae |
| Subfamily: | Myrmicinae |
| Tribe: | Pogonomyrmecini |
| Genus: | Pogonomyrmex |
| Species group: | huachucanus |
| Species: | P. huachucanus |
| Binomial name | |
| Pogonomyrmex huachucanus Wheeler, W.M., 1914
| |


This species nests in the soil, with the entrance hole often surround by a small, soil mound (up to 10-cm diameter), in fine sand soils or loam with scattered stones, or rocky soils. This species also nests under stones. Nests are usually difficult to find and may be simply small entrance holes. They are occasionally surrounded by seed hulls, which makes them easier to locate. Rarely there are a few pebbles around the entrance. Nests ttypically contain about 125 - 150 workers, although large nests may contain 400 - 500 workers. Brood is found in nests from June to July, reproductives in July and August. It forages diurnally, and uses a mixture of seeds, excrement, and dead insects as the food source and stores seeds in the nests. (Mackay and Mackay 2002)
| At a Glance | • Brachypterous Queen |
Identification
This is a member of the “Ephebomyrmex” group of species, although it serves as a ideal link between this group (“Ephebomyrmex”) and the other species in the genus. The propodeal spines are somewhat united by a carina, but the psammophore is reasonably well developed. It is a very coarsely sculptured species. The propodeal spines separate this species from others that lack spines. The rough sculpture (the area between the rugae on the head is punctate, the area between the rugae on the mesosoma is shiny, but the rugae are reticulated) separates it from many of the species which have propodeal spines. (Mackay and Mackay 2002)
Keys including this Species
Distribution
United States: southern Arizona, southern New Mexico.
Latitudinal Distribution Pattern
Latitudinal Range: 35.93822222° to 19.752917°.
| North Temperate |
North Subtropical |
Tropical | South Subtropical |
South Temperate |
- Source: AntMaps
Distribution based on Regional Taxon Lists
Nearctic Region: United States (type locality).
Neotropical Region: Mexico.
Distribution based on AntMaps
Distribution based on AntWeb specimens
Check data from AntWeb
Countries Occupied
| Number of countries occupied by this species based on AntWiki Regional Taxon Lists. In general, fewer countries occupied indicates a narrower range, while more countries indicates a more widespread species. |

|
Estimated Abundance
| Relative abundance based on number of AntMaps records per species (this species within the purple bar). Fewer records (to the left) indicates a less abundant/encountered species while more records (to the right) indicates more abundant/encountered species. |
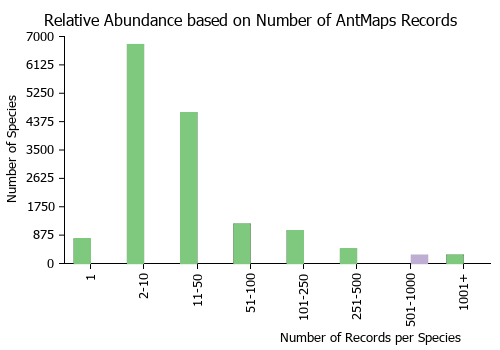
|
Habitat
Mesic or riparian areas along the edges of the Chihuahuan Desert, often in grasslands, thorn scrub, oak forests or pinyon-juniper forests, up to the edges of pine forests. (Mackay and Mackay 2002)
Biology

Worker from base of Coconino Rim, Navajo Reservation, Arizona.

Habitat: Base of Coconino Rim, Navajo Reservation, Arizona


{kind=link}
Cole (1968): Creighton (1952, p. 73) recorded Pogonomyrmex huachucanus as occurring only at rather high elevations (1,000-5,000 ft., usually), and I quite agree with him. My earlier report (Cole, 1934c, p. 400) of the ants at Needles, California (400 ft.), is indeed very badly out of line. That report, undoubtedly resulting from a misdetermination, must be declared inaccurate. I have been unable to find such representatives in my collection.
Nests of huachucanus are constructed in rather tight stony soil beneath stones, or without covering materials, in open areas exposed to the sun. Often a nest is marked only by an irregular entrance with little or no peripherally placed excavated soil. Less frequently a poorly or well-defined crater about 5 or 6 inches in diameter will have been constructed. Nest populations are small in comparison with those of members of the subgenus Pogonomyrmex and include usually no more than 125 to 150 workers. Creighton (1952, p. 72) reported that 400-500 workers may be present in exceptionally large colonies.
The workers forage singly at a slow and steady gait and return to their nests with a variable array of booty — dead arthropods, bits of excrement, and some seeds. The granivorous propensity seems not to be strongly developed in huachucanus, for seeds appear to represent only a minimal part of its diet. One sees at times a light irregular border of chaff around a nest opening. But no one yet has reported an investigation of nests other than during the summer, so it is not known whether the ants store seeds in nest chambers as winter food.
Creighton (1952, p. 73) found males in nests on July 31 (near Socorro, New Mexico) and August 5 (Carr Canyon, Huachuca Mountains, Arizona), and he took both males and dealate females from a nest on July 30 (Springerville, Arizona). I collected only males from a nest on August 3 (mouth of Madera Canyon, Santa Rita Mountains, Arizona) and both males and dealate females from nests on August 13 (16 mi. W. of Socorro, New Mexico); on August 12 (15 mi. W. of Mountainair, New Mexico) I observed in nests males and both alate (brachypterous) and dealate females.
For a number of years I have been puzzled by having found only dealate females associated with workers in nests of huachucanus although mature males were frequently present in these same nests. Creighton (p. 72) speculated about this situation. He reasoned that because there had been an hour or so of heavy rainfall just before he had excavated such a nest near Springerville, Arizona, it was conceivable that the mated and dealated females (queens) had returned to the parental nest during the wet period and had taken temporary shelter there. I (Cole, 1954a, p. 120) questioned the validity of this explanation after having found a number of nests near Mountainair, New Mexico, which contained numerous workers and males but only dealated females (in considerable numbers), and which occupied a station where no rain had fallen for many days preceding the observations. I hypothesized, therefore, that the sexes had mated in flight subsequent to which the mated and dealated females had returned to their parental nests prior to their nest-founding activities. The possibility that mating may have occurred in the nests was also suggested.
Recent collections and observations of well-established nests of huachucanus have cast a new light on this problem. Within each of three excavated nests near Socorro, New Mexico, I found workers, numerous males, a considerable number of dealate females, and a few mature alate females. The latter were scarce and represented, in my sample, a ratio of about one alate female to forty dealate females. My surprise was supreme, however, when I noted, during pointing procedure, that all of the alate females were brachypterous. An examination showed both pairs of wings to be of perfect development in shape and venation, but to have dimensions considerably less than those which would be considered as normal in the genus. The wings, fully extended in repose and with the body regions in linear alignment, reach only as far as approximately one-third the length of the dorsum of the first gastric segment. The "normal" length of flexed wings in this genus puts their tips well beyond the apex of the undistended gaster.
I believe, now, that mating must take place within the parental nest.
Mackay and Mackay (2002) - It is the only North American Pogonomyrmex species that has multiple wingless female reproductives in the nest, although it is not clear whether more than one is involved in reproduction (Pogonomyrmex anergismus, a parasitic species, may also have several females in the nest). Mating may occur in the nests, and at least some of the females are brachypterous (have small wings). Cole (1954b) suggests the females mate in flight, and return to the nest, and then subsequently leave the nest to form colonies at a later time. Workers are preyed upon by Dorymyrmex smithi. It is very docile and a pleasure to work with, and this fascinating species should be studied in detail.
Castes
Queen described as brachypterous (Johnson 2010)
Worker
   
| |
| Worker. . | Owned by Museum of Comparative Zoology. |
   
| |
| Worker. . | Owned by Museum of Comparative Zoology. |
Images from AntWeb
  
| |
| Worker. Specimen code casent0006263. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by CAS, San Francisco, CA, USA. |
Queen
   
| |
| Queen. . | Owned by Museum of Comparative Zoology. |
Images from AntWeb
   
| |
| Queen (brachypterous). Specimen code casent0173111. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by RAJC, Robert A. Johnson Collection. |
Nomenclature
The following information is derived from Barry Bolton's Online Catalogue of the Ants of the World.
- huachucanus. Pogonomyrmex huachucanus Wheeler, W.M. 1914e: 151 (w.) U.S.A. Creighton, 1952b: 79 (q.m.); Wheeler, G.C. & Wheeler, J. 1972b: 237 (l.); Taber, Cokendolpher & Francke, 1988: 51 (k.). Combination in P. (Ephebomyrmex): Cole, 1968: 157.
Type Material
- Syntype, workers, Miller Canyon, Huachuca Mts., Arizona, United States, Museum of Comparative Zoology.
Unless otherwise noted the text for the remainder of this section is reported from the publication that includes the original description.
Description
Worker
Cole (1968) - HL 1.43-1.62 mm, HW l.38-1.60 mm, CI 96.5-100.0, SL 1.02-1.14 mm, SI 68.0-74.2, EL 0.24-0.31 mm, EW 0.l9-0.22 mm, OI 16.8-19.1, WL 1.43-1.62 mm, PNL 0.36-0.46 mm, PNW 0.37-0.48 mm, PPL 0.37-0.46 mm, PPW 0.49-0.60 mm.
Mandible as shown in Pl. III, Fig. 22; all teeth rather well developed and well separated; apical tooth very broad, blunt, rather thin, considerably longer than subapical tooth; subapical tooth very broad, blunt, much broader than first basal; second basal distinctly, shorter than both first and third basals; penultimatt basal minute, very much shorter than third basal, moderately offset in two planes; mesal portion of basal mandibular margin strongly and broadly convex.
Base of antennal scape as illustrated in Pl. I, Fig. 4; shaft strongly curved in basal one-third of its length, moderately constricted hut not flattened along the bend. Basal enlargement rather prominent, its superior lobe weak, its inferior lobe strong, elongate, and subtriangular; superior declivity nearly straight, meeting the shaft at a very weak and broadly rounded angle; apex of superior lobe acute; inferior declivity long, interrupted by a well-defined point, meeting the shaft smoothly at a weak and very broad angle; basal flange prominent, extending well outward from base, its margin rather narrow, curved distad of the base, extending distinctly away from apex of superior lobe. Lateral surface of basal enlargement only weakly or not at all impressed.
Eye small, not much longer than wide, placed a little below midcenter of side of head, the distance from its anterior margin to the mandibular insertion about twice the maximum eye length. Contours of thorax, petiole, and postpetiole, in lateral view, as shown in Pl. V, Fig. 11. Anterior declivity of petiolar node broadly convex, the apex acute and bearing a prominent nipple, the dorsum flattened. Contours of petiole and post petiole, in dorsal view, as illustrated in Pl. VII, Fig. 25.
The following comprehensive description of sculpture and pilosity is quoted from Creighton (1952, pp. 76-79):
The entire upper surface of the head covered with wavy longitudinal rugae, those at the middle of the head diverging toward the occipital corners. Over most of the upper surface of the head the rugae rarely form reticulations but all the genae and the posterior quarter of the head they are distinctly reticulate. The surface between the rugae is densely and minutely granulose with the granules appearing in certain lights as tiny, crater-like rings. The upper surface of the head is dull or very feebly shining except for the anterior ends of the frontal lobes which are rather strongly shining. Antennal scapes moderately shining with prominent longitudinal rugules.
Thoracic rugae notably reticulate and not parallel on the pronotum. Those on the mesonotum and the mesopleurae also reticulate but with marc of a tendency to form roughly parallel, longitudinal ridges. These reticulate rugae turn upward and become transverse on the basal face of the epinotum. The granulate interrugal sculpture of the thorax is morc feeble than that of the head and this makes the thorax somewhat more shining than the head. Posterior face of the node of the petiole reticulo-rugose. The interrugal areas, the front of the node and the entire postpetiole densely and finely granulose, dull or at most very feebly shining. Peduncle of the petiole and the coxae with fine punctures which are more widely spaced and do not dull the shining surface. Gaster very strongly shining and with minute, widely scattered piligerous punctures. Femora strongly shining with extremely delicate reticulate sculpture. Tibiae more coarsely sculptured and not so strongly shining.
Erect hairs on the upper surface of the head stout, tapered and rather variable in length. Those on the front of the clypeus long and stout, their length equal to that of the longer hairs in the gular ammochaetae. The hairs in the latter structures rather sparse, less than a dozen present at each side of the head. Hairs of the mandibular ammochaetae much shorter and finer than those of the gula. Erect hairs of the thorax very irregular in length, abundant on the promesonotum but sparser on the epinotum, where they are largely confined to an area just behind the mesoepinotal suture. Erect hairs on the petiole and postpetiole very similar to those of the thorax. Erect gastric hairs more uniform in length than those of the thorax, their length increasing gradually from the base of the gaster to its lip. Antennal scares, coxae, femora, tibiae and tarsi with numerous, somewhat finer, erect hairs. Erect hairs [I would call them pubescence] on the antennal funiculi numerous and fine and grading to pubescence on the terminal segments.
Color: deep ferrugineous red to orange red, the mandibular borders piceous brown. Edge of first gastric segment marked with brown.
Queen
Cole (1968) - HL 1.72-1.87 mm, HW 1.75-1.82 mm, CI 96.2-101.7, SL 1.12-1.19 mm, SI 61.5-68.0, EL 0.27-0.32 mm, EW 0.20-0.22 mm, OI 15.0-18.6, WL 1.96-2.04 mm, PNL 0.54-0.63 mm, PNW 0.58-0.65 mm, PPL 0.51-0.54 mm, PPW 0.73-0.85 mm.
Eye small, weakly convex, protruding from side of head. Contours of thorax, petiole, and postpetiole as shown in Pl. IX, Fig. 4. Brachypterous; wing membrane evenly and rather densely covered with short appressed hairs, its anal margin bearing a dense fringe of rather short, delicate, whitish hairs.
I am unable to improve the following thorough and accurate account of sculpture and pilosity published by Creighton (1952, p. 79):
Cephalic sculpture very similar to that of the worker but with the longitudinal rugae at the center of the head finer and more regular. The interrugal sculpture is less pronounced than in the worker so that the entire head, especially the middle third, is more shining than that of the worker. Mandibles and antennal scapes as in the worker.
Thoracic rugae everywhere coarse and prominent except on the neck of the pronotum and the declivous face of the epinotum. The neck of the pronotum is transversely rugulose at its anterior edge. Behind this rugulose area is a band of dense, though shallow, punctures interspersed with extremely delicate rugules which extends back to the coarse, reticulate rugae on the humeral angles. Rugae of the scutum highly variable. In some specimens transverse, in others longitudinal. Rugae of the scutellum usually longitudinal. Mesothoracic sternite and episternite longitudinally rugose, the rugae turning upward on the sides of the epinotum and becoming transverse on the basal face of the epinotum. Declivous face of the epinotum with two or three feeble, widely spaced rugae. Interrugal sculpture of the thorax like that of the worker but a little more feeble so that the thorax is slightly more shining than that of the worker. Sculpture of the petiolar nodes, gaster and legs like that of the worker.
Erect hairs on the upper surface of the head, thorax, peliolar nodes and gaster slightly more numerous than those of the worker. Erect hairs elsewhere very similar to those of the worker.
Color ferrugineous red, the masticatory margin of the mandibles and the lateral edges of the dorsum of the first gastric segment banded with piceous brown.
Male
Cole (1968) - HL 0.95-1.16 mm, HW 0.97-1.16 mm, CI 100.0-102.7, SL 0.36-0.41 mm, SI 31.9-37.1, EL 0.34-0.37 mm, EW 0.24-0.27 mm, OI 31.0-35.8, WL 1.79-1.87 mm, PNL 0.27-0.34 mm, PNW 0.32-0.43 mm, PPL 0.39-0.48 mm, PPW 0.19-0.58 mm.
Head, in full-face view, as shown in Pl. II, Fig. 7; moderately narrowed between the eyes and mandibular insertions, the constricted portions broadly convex and notably convergent anteriorly; transverse occipital keel strongly developed; frontal area large, triangular, deeply impressed. Head, in lateral view, as illustrated in Pl. II, Fig. 8; occipital corner acute; eye moderately large, its anterior border separated from the mandibular insertion by a distance approximately equal to the maximum eye length. Mandible as shown in Pl. VIII, Fig. 17; blade large, broad apically; masticatory margin oblique, bearing 6 well-spaced teeth; basal margin broadly convex. Segments 4-9, inclusive, of flagellum incrassate mesally. Antennal scape more than one-half as long as length of first two flagellar segments combined, it and the first flagellar segment subequal in length, the widest portions of both subequal.
Epinotal armature notably compressed laterally. Hair pattern of wing membrane as in the female. Contour of petiole and postpetiole, in dorsal view, as illustrated in Pl. XII, Fig. 16.
Genitalia, in dorsal view, as shown in Pl. X, Fig. 17; terminal lobe of left paramere, in inner lateral view, as in Pl. XI, Fig. 17; contour of left volsella, in inner lateral view, as in Pl. XII, Fig. 7. Aedeagus (Pl. X, Fig. 18) with a twist which brings the proximal half of the serrate ventral margin strongly mesad and produces a broadly triangular portion with a prominent, sharp mesoproximal apex; denticles on ventral margin small but rather pronounced, clearly visible at a magnification of 90X.
The following description of sculpture and pilosity is quoted from Creighton (1952, p. 80):
Upper surface of the head with fine, wavy, longitudinal fugae over most of its middle third. These rugae form few reticulations. The rugae of the lateral portions of the head, the occipital angles, the entire occipital border and the gula, notably reticulate. The surface between the rugae dull and densely punctured, the punctures appearing as tiny craters under high magnification. Frontal groove, frontal area and the clypeus slightly shining. Mandibles longitudinally striate and a little more shining than the clypeus.
Entire thorax feebly shining to dull with punctato-rugose sculpture, the punctures very dense. Rugae on the pronotum, mesothoracic sternite and the episternite feeble, often replaced by punctures on the front part of the segments. Scutum, paraptera and the scutellum with moderately developed longitudinal, reticulate rugae in addition to the dense punctures. Metathoracic sternite with coarse longitudinal rugae which turn upward on the sides of the epinotum. Basal face of the epinotum with coarse, reticulate rugae. Rugae on the declivous face of the epinotum transverse. Anterior peduncle of the petiole finely punctate and more shining ul,m the remainder of the petiole. The anterior face of the node of the petiole densely and more coarsely punctate. Crest and rear face of the node and the posterior peduncle with reticulate rugae in addition to the punctures. Postpetiole densely punctate but not rugose. Abdomen smooth and shining with extremely fine, scattered, piligerous punctures. Coxal joints, antennal scapes, fore femora and tarsi delicately reticulo-punctate. Middle and hind femora with similar but feebler sculpture.
Erect hairs of moderate length, yellow-brown, evenly curved, tapered and well-separated. Abundant on the dorsum of the thorax, except on the basal face of the epinotum, where they are restricted to the angle between the basal and the declivous faces. Erect hairs on the head about equally numerous on the upper and lower surfaces. Erect hairs on the petiolar nodes notably sparser than those of the thorax. Erect hairs on the gaster very fine, much shorter than those of the thorax and largely limited to the rear edge of each segment. The remaining gastric hairs very short and usually fully appressed, rarely a few of them suberect. Antennal scapes and funiculi with dense, short, pubescence.
Color: head, thorax, petiolar nodes and femora piceous black. Antennae, toothed border of the mandibles, tibiae and tarsi blackish brown. Gaster clear, golden yellow.
Wings hyaline, iridescent, the veins and stigma brown in some specimens and white in others. The wings evenly covered with small scattered hairs.
I should like to add to Creighton's description of the male the following characteristics: hairs are absent from the antennal scapes and from the venter of the petiole and its peduncle; the eyes bear very short and rather sparse hairs; the hairs on the legs are quite sparse and, for the most part, coarse and decumbent; the wing membrane is not only pilose, but also bears on chiefly its anal margin a prominent fringe of hairs; and the color of the gaster vanes from a clear, golden yellow to a notably infuscated yellow.
Karyotype
- See additional details at the Ant Chromosome Database.
 Explore: Show all Karyotype data or Search these data. See also a list of all data tables or learn how data is managed.
Explore: Show all Karyotype data or Search these data. See also a list of all data tables or learn how data is managed.
- 2n = 36 (USA) (Taber et al., 1988).
References
- Alatorre-Bracamontes, C.E., Vásquez-Bolaños, M. 2010. Lista comentada de las hormigas (Hymenoptera: Formicidae) del norte de México. Dugesiana 17(1): 9-36.
- Cole, A. C., Jr. 1968. Pogonomyrmex harvester ants. A study of the genus in North America. Knoxville, Tenn.: University of Tennessee Press, x + 222 pp. (page 157, Combination in P. (Ephebomyrmex))
- Creighton, W. S. 1952b. Studies on Arizona ants (3). The habits of Pogonomyrmex huachucanus Wheeler and a description of the sexual castes. Psyche (Camb.) 59: 71-81. (page 79, queen, male described)
- Jansen, G., Savolainen, R. 2010. Molecular phylogeny of the ant tribe Myrmicini (Hymenoptera: Formicidae). Zoological Journal of the Linnean Society 160(3), 482–495 (doi:10.1111/j.1096-3642.2009.00604.x).
- Johnson RA. 2010: Independent colony founding by ergatoid queens in the ant genus Pogonomyrmex: queen foraging provides an alternative to dependent colony founding. Insectes Sociaux 57: 169–176.
- Johnson, R.A. 2010. Independent colony founding by ergatoid queens in the ant genus Pogonomyrmex: queen foraging provides an alternative to dependent colony founding. Insectes Sociaux 57, 169–176 (doi:10.1007/s00040-010-0065-6).
- Mackay, W. P. and E. Mackay. 2002. The ants of New Mexico (Hymenoptera: Formicidae). Edwin Mellen Press, Lewiston, NY.
- Taber, S. W.; Cokendolpher, J. C.; Francke, O. F. 1988. Karyological study of North American Pogonomyrmex (Hymenoptera: Formicidae). Insectes Soc. 35: 47-60 (page 51, karyotype described)
- Wheeler, G. C.; Wheeler, J. 1972b. Ant larvae of the subfamily Myrmicinae: second supplement on the tribes Myrmicini and Pheidolini. J. Ga. Entomol. Soc. 7: 233-246 (page 237, larva described)
- Wheeler, W. M. 1914f. New and little known harvesting ants of the genus Pogonomyrmex. Psyche (Camb.) 21: 149-157. (page 151, worker described)
References based on Global Ant Biodiversity Informatics
- Cole A. C., Jr. 1937. An annotated list of the ants of Arizona (Hym.: Formicidae). [concl.]. Entomological News 48: 134-140.
- Cover S. P., and R. A. Johnson. 20011. Checklist of Arizona Ants. Downloaded on January 7th at http://www.asu.edu/clas/sirgtools/AZants-2011%20updatev2.pdf
- Creighton W. S. 1952. Studies on Arizona ants (3). The habits of Pogonomyrmex huachucanus Wheeler and a description of the sexual castes. Psyche (Cambridge) 59: 71-81.
- Dattilo W. et al. 2019. MEXICO ANTS: incidence and abundance along the Nearctic-Neotropical interface. Ecology https://doi.org/10.1002/ecy.2944
- Fernandes, P.R. XXXX. Los hormigas del suelo en Mexico: Diversidad, distribucion e importancia (Hymenoptera: Formicidae).
- Johnson R. A., and C. S. Moreau. 2016. A new ant genus from southern Argentina and southern Chile, Patagonomyrmex (Hymenoptera: Formicidae). Zootaxa 4139: 1-31.
- Johnson R. Personnal Database. Accessed on February 5th 2014 at http://www.asu.edu/clas/sirgtools/resources.htm
- Mackay, W.P., E.E. Mackay, J.F. Perez Dominguez, L.I. Valdez Sanchez and P.V. Orozco. 1985. Las hormigas del estado de Chihuahua Mexico: El genero Pogonomyrmex (Hymenoptera: Formicidae) . Sociobiology 11(1):39-54
- Mackay W. P., and E. E. Mackay. 2002. The ants of New Mexico (Hymenoptera: Formicidae). Lewiston, New York: Edwin Mellen Press, 400 pp.
- Mallis A. 1941. A list of the ants of California with notes on their habits and distribution. Bulletin of the Southern California Academy of Sciences 40: 61-100.
- Parker, J.D. and S.W. Rissing. 2002. Molecular Evidence for the Origin of Workerless Social Parasites in the Ant Genus Pogonomyrmex. Evolution 56(10):2017-2028
- Strehl, C.-P. and J. Gadau. 2004. Cladistic Analysis of Paleo-Island Populations of the Florida Harvester Ant (Hymenoptera: Formicidae) Based upon Divergence of Mitochondrial DNA Sequences. The Florida Entomologist 87(4):576-581
- Taber S. W., J. C. Cokendolpher, and O. F. Francke. 1988. Karyological study of North American Pogonomyrmex (Hymenoptera: Formicidae). Insectes Soc. 35: 47-60.
- Vásquez-Bolaños M. 2011. Lista de especies de hormigas (Hymenoptera: Formicidae) para México. Dugesiana 18: 95-133