Linepithema iniquum
| Linepithema iniquum | |
|---|---|

| |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Arthropoda |
| Class: | Insecta |
| Order: | Hymenoptera |
| Family: | Formicidae |
| Subfamily: | Dolichoderinae |
| Tribe: | Leptomyrmecini |
| Genus: | Linepithema |
| Species: | L. iniquum |
| Binomial name | |
| Linepithema iniquum (Mayr, 1870)
| |
| Synonyms | |
| |


{kind=link}
A widely ranging arboreal species.
| At a Glance | • Invasive |
Identification
Wild (2007) - Worker Mesonotal dorsum with deep, step-like mesal impression; dorsum of head posterior of clypeus with > 5 erect setae; mesopleura and metapleural bulla lacking pubescence and strongly shining.
L. iniquum exhibits considerable variation among the worker caste in size, color, propodeal form, pilosity, and pubescence
Workers of the closely-related Linepithema dispertitum are structurally similar but the mesonotal impression is normally less well developed, the scapes often somewhat shorter, and in most populations have fewer than 4 standing setae on the dorsum of the head posterior to the clypeus. In the region where the two species co-occur, from Guatemala to Panamá, L. iniquum often shows a reduction in the development of the mesonotal impression such that it occasionally approaches the condition of L. dispertitum. However, the two species in this region can usually be separated by differences in pilosity and pubescence. Linepithema dispertitum, while pilose in parts of northern Mexico, lacks the extensive pilosity of L. iniquum in southern Central America, having four or fewer erect setae on the head, and always bearing dense pubescence on gastric tergites 1 and 2. Linepithema dispertitum also shows dense gastric pubescence on gastric tergites 1–2 (sometimes dilute on 3–4), while L. iniquum collections from this region nearly always have sparse gastric pubescence on tergites 2–4. Additionally, L. dispertitum frequently nests in soil or rotting wood, while L. iniquum is normally arboreal. Males of Linepithema leucomelas are nearly indistinguishable from L. iniquum males, but the petiolar node of L. leucomelas males is much lighter in color than the mesosomal dorsum.
Male Forewing with 1 submarginal cell; petiolar node bearing a relatively low, rounded scale; propodeum with posterior face straight to convex; petiolar node concolorous with mesosoma; eye separated from posterolateral margin of clypeus by a distance less than or equal to width of antennal scape.
Keys including this Species
- Clave para Linepithema en Colombia
- Key to Linepithema males
- Key to Linepithema of Columbia
- Key to Linepithema workers
Distribution
Central America, Jamaica, Puerto Rico, the Lesser Antilles, and the northern coast of South America south along the Andes to Paraguay and southeastern Brazil. Incidental worldwide.
Latitudinal Distribution Pattern
Latitudinal Range: 25.68015° to -28.93333333°.
| North Temperate |
North Subtropical |
Tropical | South Subtropical |
South Temperate |
- Source: AntMaps
Distribution based on Regional Taxon Lists
Neotropical Region: Argentina, Brazil, Colombia (type locality), Costa Rica, Ecuador, El Salvador, French Guiana, Greater Antilles, Grenada, Guatemala, Guyana, Lesser Antilles, Martinique, Mexico, Panama, Paraguay, Puerto Rico, Suriname, Venezuela.
Palaearctic Region: United Kingdom of Great Britain and Northern Ireland.
Distribution based on AntMaps
Distribution based on AntWeb specimens
Check data from AntWeb
Countries Occupied
| Number of countries occupied by this species based on AntWiki Regional Taxon Lists. In general, fewer countries occupied indicates a narrower range, while more countries indicates a more widespread species. |

|
Estimated Abundance
| Relative abundance based on number of AntMaps records per species (this species within the purple bar). Fewer records (to the left) indicates a less abundant/encountered species while more records (to the right) indicates more abundant/encountered species. |
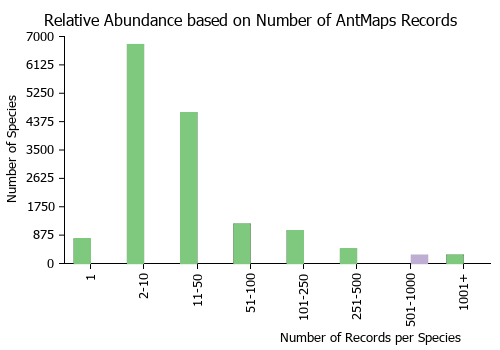
|
Biology
Wild (2007) - Linepithema iniquum is primarily an arboreal ant. Thirteen nest records are from dead branches or dead wood inside living trees, four are from dead twigs or vines, two records are from the base of a bromeliad, one from a bamboo sheath, two from rotting wood on the ground, and nine nests intercepted in ports-of-entry around the world have been in orchids. Wheeler (1908: 153) reports the ant in Puerto Rico (as Iridomyrmex melleus) nesting in hollow twigs, in leaf petioles of plantain, and in “friable carton (earth mixed with vegetable debris) on the under sides of the huge reniform leaves [of Coccoloba rugosa (Ortegon)],” and in Dominica “‘under and in dead stalks of bananas and plantains’”(Wheeler 1913b: 242). On St. John Island, Pressick and Herbst (1973) report L. iniquum nesting in twigs and in logs in moist forest and grassy field habitats. Leuderwaldt (1926) reports two nests in bamboo (as I. iniquus), one under bark (as I. iniquus var. succinea), and one in a dry branch (as I. iniquus var. succinea) in southern Brazil. Linepithema iniquum has been collected rarely in leaf-litter surveys, but these likely reflect incidental ground foragers.
Linepithema iniquum has been collected from sea level to over 2000 meters in elevation. This species is exclusively montane in Central America and northern South America, and is found at varying elevations in the Caribbean, southern Brazil, and Paraguay. 25 museum records are from human disturbed habitats such as 2nd growth forest edges, roadsides, orchards, and pastures. Four are from tropical humid montane forest, seven from primary Atlantic forest in Brazil and Paraguay, three Paraguayan records are Malaise trap samples from low inundated forest, and one Paraguayan record is from an inundated grassland. In Ecuador this species can be locally abundant along roadsides, in pastures, and in Psidium guayaba orchards, and in Puerto Rico L. iniquum is among the most abundant ants in the mountains (Wheeler 1908, Wild, pers obs).
Linepithema iniquum is probably polydomous, as individual arboreal nests frequently contain no queens (Wild, pers obs). Of five full nest excavations conducted by the author in Ecuador and Puerto Rico that found dealate queens, four colonies had a single dealate queen and one had two dealate queens. This pattern suggests that L. iniquum is monogynous to weakly polygynous, although confirmation will require molecular genetic data. Male and female alates have been observed in nests in Brazil from October to April, in Ecuador in August and December, in Costa Rica in November, and in Puerto Rico year round. Both male and female alates have been attracted to lights in November in Puerto Rico.
In Puerto Rico this species has been observed tending Coccus and Saissetia scale on coffee (Smith 1942: 22), tending pseudococcids on Cecropia (Wild, pers. obs), and visiting extra floral nectaries (Wild, pers. obs). There is one observation of aphid-tending in Ecuador (Wild, pers. obs). Smith (1929) and Wheeler (1929) describe the behavior of introduced L. iniquum in North American greenhouses in Illinois and Massachusetts, respectively, and observe the ant tending scale, visiting flowers, and nesting in and under soil pots.
Like its better-known congener Linepithema humile, L. iniquum is carried around the world with human commerce. More than a dozen museum records of this species are intercepts at various ports-of-entry and quarantine in the United States and Europe, usually carried with epiphytes. Unlike L. humile, this species apparently has not been successful in establishing outdoor populations in spite of the opportunity to do so, although there are a few records of this ant persisting in greenhouses in the temperate zone (Wheeler 1929, Creighton 1950).
Escarraga and Guerrero (2016) - Columbia Linepithema iniquum is an arboreal ant that prefers high altitude places. In some localities, this species was collected in coffee crops and trees. In one collection from the Sierra Nevada de Santa Marta, sector San Pedro de la Sierra, this species was found nesting inside a stick on the leaf litter along with an unidentified Pheidole. In the stick, larvae and pupae of both species were found, in addition to L. iniquum males. In the laboratory, the L. iniquum workers seemed to feign death (thanatosis) when threatened. Food chambers were observed, containing the heads and bodies of insects such as bees (Halictidae) and flies (Diptera).
Rosumek (2017) assessed trophic resource use and period of activity within the ant community in a southern Brazil Atlantic forest. He found a number of Linepithema, reporting "More is known about Linepithema iniquum, mainly because it also appears as an exotic indoor species in North America and Europe. A few instances of honeydew and extra-floral nectaries use exist in the literature (Wheeler, 1929; Wild, 2007; Schmid et al., 2010) and Smith (1929) described it collecting arthropods, although without specifying if that meant scavenging or predation. Wild (2007) describes it as a primary arboreal ant, but clearly it also forages on the ground. None of the Linepithema species sampled in this study showed statistically significant preferences, due to low number of records and use of several resources. Linepithema micans and Linepithema pulex seem to use resources more broadly than Linepithema iniquum, which might descend to the ground mostly to scavenge animal resources. Linepithema iniquum was found to mostly be a diurnal forager.
Castes
Worker
     
| |
| . | |
Nomenclature
The following information is derived from Barry Bolton's Online Catalogue of the Ants of the World.
- iniquum. Hypoclinea iniqua Mayr, 1870a: 392, fig. 10 (w.) COLOMBIA. Forel, 1908c: 395 (m.). Combination in Iridomyrmex: Emery, 1890b: 56; in Linepithema: Shattuck, 1992a: 16. Senior synonym of bicolor, dominicensis, fuscescens, melleus, nigellus, succineus and material of the unavailable name paranaensis referred here: Wild, 2007a: 67.
- nigellus. Iridomyrmex iniquus var. nigellus Emery, 1890b: 56 (w.) COSTA RICA. [Also described as new by Emery, 1894k: 61.] Combination in Linepithema: Shattuck, 1992a: 16. Subspecies of dispertitum: Forel, 1901c: 134; Forel, 1912h: 47; of iniquum: Creighton, 1950a: 342. Junior synonym of iniquum: Wild, 2007a: 67.
- fuscescens. Iridomyrmex melleus var. fuscescens Wheeler, W.M. 1908a: 153 (w.) PUERTO RICO. Combination in Linepithema: Shattuck, 1992a: 16. Junior synonym of iniquum: Wild, 2007a: 68.
- melleus. Iridomyrmex melleus Wheeler, W.M. 1908a: 151, pl. 12, figs. 15, 16 (w.q.m.) PUERTO RICO. Wheeler, G.C. & Wheeler, J. 1974b: 397 (l.). Combination in Linepithema: Shattuck, 1992a: 16. Junior synonym of iniquum: Wild, 2007a: 68.
- succineus. Iridomyrmex melleus subsp. succineus Forel, 1908c: 396 (w.) BRAZIL. Combination in Linepithema: Shattuck, 1992a: 16. Subspecies of melleum: Santschi, 1929d: 305; of iniquum: Forel, 1912h: 47; Kempf, 1972a: 125. Junior synonym of iniquum: Wild, 2007a: 68.
- bicolor. Iridomyrmex iniquus var. bicolor Forel, 1912h: 47 (w.) MARTINIQUE I. Combination in Linepithema: Shattuck, 1992a: 16. Junior synonym of iniquum: Wild, 2007a: 68.
- dominicensis. Iridomyrmex melleus var. dominicensis Wheeler, W.M. 1913d: 242 (w.) DOMINICA. Combination in Linepithema: Shattuck, 1992a: 16. Junior synonym of iniquum: Wild, 2007a: 68.
Unless otherwise noted the text for the remainder of this section is reported from the publication that includes the original description.
Wild (2007):
L. iniquum exhibits considerable variation among the worker caste in size, color, propodeal form, pilosity, and pubescence, which has lead to a proliferation of names. I find little justification in retaining the multiplicity of names associated with L. iniquum and Linepithema melleum. Most of these names describe local color forms, and these forms inevitably lose their distinctness with the examination of sufficient quantity of material. Much of the variation is allopatric. The few instances of sympatric variation are normally with respect to only one character, and usually the character is present in an intermediate state either in the same location or within specimens collected within few hundred kilometers. Males are remarkably similar in form and in genital structure across the range, differing primarily in color and more rarely in mesonotal pilosity. The arboreal nesting habit is also apparently uniform across the range.
Specimens from the eastern Andean slopes in Ecuador, Peru, and parts of Colombia, as well as from Jamaica and some high elevations in Central America, are nearly always medium brown to black in color and have sparse pubescence on gastric tergites 2–4. Mayr’s Hypoclinea iniqua holotype from Bogota is damaged, with only a gaster and a few tarsi remaining, but it unambiguously belongs to this form as the gaster has the distinctive sparse pubescence and dark coloration. Costa Rican specimens, including Emery’s types of I. iniquus nigellus, are often more robust than most other L. iniquum, with a less pronounced mesonotal impression, but these are otherwise similar to other dark sparsely pubescent forms. Specimens from the western Andean slopes from Ecuador north through Calí, Colombia, as well as from Guyana north through the Lesser Antilles to Puerto Rico, have moderate to dense pubescence on all gastric tergites. These pubescent forms are variable in color, from yellow to dark brown, without much geographic structure to the color variation. Puerto Rican specimens are most commonly pale yellow, as in Wheeler’s types of I. melleus, but there is continuous variation to bicolored or to a medium brown, as in Wheeler’s I. melleus fuscecens. Wheeler’s I. melleus dominicensis types from Dominica appear indistinguishable from paler specimens found in Puerto Rico. Venezuelan and northern Colombian material may be of either pubescence form or intermediate, with both forms recorded in sympatry in Valle de Cauca, Colombia. Forel’s I. iniquus bicolor types from Martinique are sparsely pubescent and sharply bicolored with a yellow head and mesosoma and a dark gaster.
Atlantic forest material from Brazil presents a more confusing array of variation, as specimens can be small to large, with scapes of varying length, pale yellow to black or bicolored, and sparsely to densely pubescent, with little apparent geographic pattern to the variation and occasional local sympatry of more than one form. A dark densely pubescent form, a yellow sparsely pubescent form, and intermediates have been recorded in sympatry at Nova Teutônia, Santa Catarina. A dark sparsely pubescent form, a yellow densely pubescent form, and a bicolored form are all recorded from Rio de Janeiro and São Paulo states, sometimes in local sympatry. Many specimens from Paraná state are bicolored. Paraguayan material is dark brown, sparsely pubescent, with relatively small eyes (50–60 ommatidia).
Forel’s types of I. iniquus succineus from São Paulo, as well as several other collections from São Paulo and Rio de Janeiro states, are distinct in the combination of large size, dense gastric pubescence, and yellow-orange coloration. Further study into the ecology and genetics of this form may reveal that Linepithema succineum merits elevation to specific status (conceivably rendering the remainder of L. iniquum paraphyletic). However, given that each of the character states that in combination define this form can be found individually or in other combinations among other southern Brazilian collections of L. iniquum, I find it preferable to retain L. succineum as a synonym of L. iniquum.
Description
Worker
(n = 100) HL 0.55–0.74, HW 0.46–0.70, MFC 0.13–0.20, SL 0.55–0.78, FL 0.45–0.69, LHT 0.44–0.78, PW 0.29–0.43, ES 1.06–2.03, SI 95–129, CI 84–97, CDI 24–31, OI 19–29.
Head in full face view nearly oval in shape, varying from relatively narrow to relatively broad (CI 84–97), lateral margins convex, posterior margin convex to straight. Compound eyes of moderate size (OI 19–29), comprised of 50–105 (mean = 67) facets. Antennal scapes long (SI 95–129), slightly shorter to slightly longer than head length. In full face view, scapes in repose exceeding posterior margin of head by a length greater than the length of the first funicular segment. Frontal carinae moderately spaced (CDI 24–31). Maxillary palps long, more than ½ HL, ultimate segment (segment six) longer than segment 2.
Mesosoma in lateral view relatively slender with dorsal profile comprised of three distinct convexities: 1) pronotum + anterior mesonotum, 2) posterior mesonotum, 3) propodeum. Mesonotum with a deep, step-like mesal impression. Mesothorax at constriction usually narrower than widest diameter of fore coxa. Metanotal groove deeply impressed. Propodeum raised and varying in profile from evenly rounded to somewhat flattened dorsally. Metapleural bulla relatively swollen and protruding.
Petiolar scale inclined anteriorly with a relatively blunt node, in lateral view falling short of propodeal spiracle.
Cephalic dorsum (excluding clypeus) bearing 5–13 (mean = 7.7) erect setae, usually arranged in two parallel longitudinal rows extending from the frontal carinae towards the posterior margin. Pronotum bearing 0–7 (mean = 3.4) erect setae, anterior setae usually much longer than posterior setae when posterior setae present. Mesonotum without erect setae. Gastric tergite 1 (= abdominal tergite 3) bearing 3–7 (mean = 4.6) erect setae, tergite 2 with 1–14 (mean = 10.7) erect setae, tergite 3 with 4–11 (mean = 7.0) erect setae. Venter of metasoma with scattered erect setae.
Surface of head and mesosomal dorsum smooth and relatively shining. Pubescence variable, sparse to dense on cephalic dorsum, mesosoma, and gaster, usually more dense on cephalic dorsum than sides of head, and on anterior than posterior gastric tergites. Mesopleura and metapleural bulla always without pubescence, surface glabrous.
Color variable; entire body and appendages pale yellow to brown or black, sometimes bicolored with gaster darker than head, mesosoma, and appendages.
Queen
(n = 6) HL 0.78–0.82, HW 0.73–0.77, SL 0.72–0.76, FL 0.70–0.76, LHT 0.75–0.82, EL 0.23–0.25, MML 1.44–1.63, WL 4.11–4.92, CI 92–97, SI 94–101, OI 30–31, WI 29–30, FI 45–50.
Moderately small species (MML < 1.7). Head longer than broad in full face view (CI 92–97), posterior margin straight to slightly convex. Eyes small (OI 30–31). Ocelli small. Antennal scapes moderate to long (SI 94–101), in full face view scapes in repose surpassing posterior margin by a length greater than length of first funicular segment.
Forewings moderately short relative to mesosomal length (WI 29–30). Forewings with Rs+M somewhat longer than M.f2. Legs moderately long relative to mesosomal length (FI 45–50).
Dorsum of mesosoma and metasoma with abundant standing setae. Mesoscutum bearing more than 10 standing setae. Body color variable, from testaceous to piceous.
Male
(n = 5) HL 0.43–0.56, HW 0.41–0.52, SL 0.10–0.15, FL 0.46–0.63, LHT 0.41–0.56, EL 0.19–0.23, MML 0.78–1.01, WL 1.93–2.66, PH 0.17–0.21, CI 91–99, SI 24–26, OI 40–45, WI 23–28, FI 57–62.
Head relatively narrow to about as broad as long in full face view (CI 91–99). Eyes of moderate size (OI 40–45), occupying much of anterolateral surface of head anterior of midpoint and separated from posterolateral clypeal margin by a length less than or equal to width of antennal scape. Ocelli small and in full frontal view emerging only slightly above adjoining posterolateral margins. Antennal scape of moderate length (SI 24–26), about 55–65% length of 3rd antennal segment. Anterior clypeal margin convex medially. Mandibles variable; most commonly mandibles moderately small, masticatory margin slightly longer than inner margin, inner margin and exterior lateral margin parallel to slightly diverging, apical tooth enlarged and followed by a series of 8–12 denticles. More rarely mandibles worker-like, masticatory margin broad and much longer than inner margin, inner margin and exterior lateral margin diverging, apical 2–3 teeth enlarged and followed by a series of 14–18 denticles.
Mesosoma not well developed and subequal in length to metasoma. Mesoscutum not greatly enlarged, not projecting forward over pronotum. Propodeum in lateral view not well developed and not overhanging petiole, posterior face slightly convex and rounding gradually into dorsal face. Forewings of moderate length (WI 23–28) and bearing one submarginal cell. Wings transparent, with yellow, pale brown, or medium brown wing veins and stigma. Legs of moderate length relative to mesosoma (FI 57–62).
Petiolar node bearing a low, rounded scale, node height shorter than node length. Venter of node bearing a convex posteriorly-pointing process. Gaster ovoid in dorsal view, about 2 times as long as broad. Gonostylus produced as triangular pilose lobe. Volsella with cuspis present, digitus short and downturned distally.
Dorsal surfaces of body with erect setae sparse, mesoscutum with 0–10 fine erect to suberect setae. Venter of gaster with scattered setae. Pubescence dense on body and appendages, becoming sparse only on medial propodeal dorsum. Sculpture on head and mesoscutum not well developed, surface shining through pubescence.
Head, mesosoma, and metasoma testaceous to dark brown. Mandibles, antennae, and legs usually lighter than head and mesosoma.
Type Material
Holotype worker [damaged, only gaster and tarsi remaining] Distrito Capital, Santa Fe de Bogota, Colombia, Lindig Naturhistorisches Museum Wien, Vienna, examined.
References
- Cantone S. 2017. Winged Ants, The Male, Dichotomous key to genera of winged male ants in the World, Behavioral ecology of mating flight (self-published).
- Dekoninck, W., Wauters, N., Delsinne, T. 2019. Capitulo 35. Hormigas invasoras en Colombia. Hormigas de Colombia.
- Emery, C. 1890b. Voyage de M. E. Simon au Venezuela (Décembre 1887 - Avril 1888). Formicides. Ann. Soc. Entomol. Fr. (6)(10): 55-76 (page 56, Combination in Iridomyrmex)
- Escarraga, M., Guerrero, R.J. 2016. The ant genus Linepithema (Formicidae Dolichoderinae) in Colombia. Zootaxa 4208: 446–458 (DOI 10.11646/zootaxa.4208.5.3).
- Forel, A. 1908h. Ameisen aus Sao Paulo (Brasilien), Paraguay etc. gesammelt von Prof. Herm. v. Ihering, Dr. Lutz, Dr. Fiebrig, etc. Verh. K-K. Zool.-Bot. Ges. Wien 58: 340-418 (page 395, male described)
- Franco, W., Ladino, N., Delabie, J.H.C., Dejean, A., Orivel, J., Fichaux, M., Groc, S., Leponce, M., Feitosa, R.M. 2019. First checklist of the ants (Hymenoptera: Formicidae) of French Guiana. Zootaxa 4674, 509–543 (doi:10.11646/zootaxa.4674.5.2).
- Hamer, M.T., Cocks, L.R. 2020. Linepithema iniquum (Mayr) (Hymenoptera: Fromicidae) found at the National Botanic Garden of Wales. British Journal of Entomology & Natural History 33:71-75.
- Ladino, N., Feitosa, R.M. 2022. Ants (Hymenoptera: Formicidae) of the Parque Estadual São Camilo, an isolated Atlantic Forest remnant in western Paraná, Brazil. ZOOLOGIA 39: e22001 (doi:10.1590/S1984-4689.v39.e22001).
- Mayr, G. 1870a. Formicidae novogranadenses. Sitzungsber. Kais. Akad. Wiss. Wien Math.-Naturwiss. Cl. Abt. I 61: 370-417 (page 392, fig. 10 worker described)
- Rosumek, F. B. 2017. Natural History of Ants: What We (do not) Know about Trophic and Temporal Niches of Neotropical Species. Sociobiology. 64:244-255. doi:10.13102/sociobiology.v64i3.1623
- Shattuck, S. O. 1992a. Review of the dolichoderine ant genus Iridomyrmex Mayr with descriptions of three new genera (Hymenoptera: Formicidae). J. Aust. Entomol. Soc. 31: 13-18 (page 16, Combination in Linepithema)
- Varela-Hernández, F., Medel-Zosayas, B., Martínez-Luque, E.O., Jones, R.W., De la Mora, A. 2020. Biodiversity in central Mexico: Assessment of ants in a convergent region. Southwestern Entomologist 454: 673-686.
- Wetterer, J.K. 2021. Ants (Hymenoptera, Formicidae) of St. Vincent, West Indies. Sociobiology 68, e6725 (doi:10.13102/sociobiology.v68i2.6725).
- Wheeler, W. M. 1929. Two Neotropical ants established in the United States. Psyche. 36:89-90.
- Wild, A. L. 2007a. Taxonomic revision of the ant genus Linepithema (Hymenoptera: Formicidae). University of California Publications in Entomology. 126:1-159.
- Wild, A.L. 2004. Taxonomy and distribution of the Argentine ant, Linepithema humile (Hymenoptera: Formicidae). Annals of the Entomological Society of America 97, 1204-1215.
References based on Global Ant Biodiversity Informatics
- Baroni Urbani C. 1977. Katalog der Typen von Formicidae (Hymenoptera) der Sammlung des Naturhistorischen Museums Basel (2. Teil). Mitt. Entomol. Ges. Basel (n.s.) 27: 61-102.
- Borgmeier T. 1923. Catalogo systematico e synonymico das formigas do Brasil. 1 parte. Subfam. Dorylinae, Cerapachyinae, Ponerinae, Dolichoderinae. Archivos do Museu Nacional (Rio de Janeiro) 24: 33-103.
- Clemes Cardoso D., and J. H. Schoereder. 2014. Biotic and abiotic factors shaping ant (Hymenoptera: Formicidae) assemblages in Brazilian coastal sand dunes: the case of restinga in Santa Catarina. Florida Entomologist 97(4): 1443-1450.
- Clemes Cardoso D., and M. Passos Cristiano. 2010. Myrmecofauna of the Southern Catarinense Restinga sandy coastal plain: new records of species occurrence for the state of Santa Catarina and Brazil. Sociobiology 55(1b): 229-239.
- Culver D. C. 1974. Species packing in Caribbean and North Temperate ant communities. Ecology 55(5): 974-988.
- Drose W., L. R. Podgaiski, C. Fagundes Dias, M. de Souza Mendonca. 2019. Local and regional drivers of ant communities in forest-grassland ecotones in South Brazil: A taxonomic and phylogenetic approach. Plos ONE 14(4): e0215310.
- Emery C. 1890. Studii sulle formiche della fauna neotropica. Bull. Soc. Entomol. Ital. 22: 38-8
- Emery C. 1894. Estudios sobre las hormigas de Costa Rica. Anales del Museo Nacional de Costa Rica 1888-1889: 45-64.
- Emery C. 1913. Hymenoptera. Fam. Formicidae. Subfam. Dolichoderinae. Genera Insectorum 137: 1-50.
- Favretto M. A., E. Bortolon dos Santos, and C. J. Geuster. 2013. Entomofauna from West of Santa Catarina State, South of Brazil. EntomoBrasilis 6 (1): 42-63.
- Fernandes T. T., R. R. Silva, D. Rodrigues de Souza-Campana, O. Guilherme Morais da Silva, and M. Santina de Castro Morini. 2019. Winged ants (Hymenoptera: Formicidae) presence in twigs on the leaf litter of Atlantic Forest. Biota Neotropica 19(3): http://dx.doi.org/10.1590/1676-0611-bn-2018-0694
- Fernandes T. T., R. Rosa da Silva, D. Rodrigues de Souza, N. Araujo, and M. Santina de Castro Morini. 2012. Undecomposed Twigs in the Leaf Litter as Nest-Building Resources for Ants (Hymenoptera: Formicidae) in Areas of the Atlantic Forest in the Southeastern Region of Brazil. Psyche doi:10.1155/2012/896473
- Fernandes T. T., W. Dattilo, R. R. Silva, P. Luna, C. M. Oliveira, and M. Santina de Castro Morini. 2019. Ant occupation of twigs in the leaf litter of the Atlantic Forest: influence of the environment and external twig structure. Tropical Conservation Science 12: 1-9.
- Fernández F., E. E. Palacio, W. P. Mackay, and E. S. MacKay. 1996. Introducción al estudio de las hormigas (Hymenoptera: Formicidae) de Colombia. Pp. 349-412 in: Andrade M. G., G. Amat García, and F. Fernández. (eds.) 1996. Insectos de Colombia. Estudios escogidos. Bogotá: Academia Colombiana de Ciencias Exactas, Físicas y Naturales, 541 pp
- Fernández, F. and S. Sendoya. 2004. Lista de las hormigas neotropicales. Biota Colombiana Volume 5, Number 1.
- Figueiredo C. J. de, R. R. da Silva, C. de Bortoli Munhae, and M. S. de Castro Morini. 2013. Ant fauna (Hymenoptera: Formicidae) attracted to underground traps in Atlantic Forest. Biota Neotrop 13(1): 176-182
- Forel A. 1908. Ameisen aus Sao Paulo (Brasilien), Paraguay etc. gesammelt von Prof. Herm. v. Ihering, Dr. Lutz, Dr. Fiebrig, etc. Verhandlungen der Kaiserlich-Königlichen Zoologisch-Botanischen Gesellschaft in Wien 58: 340-418.
- Forel A. 1912. Formicides néotropiques. Part V. 4me sous-famille Dolichoderinae Forel. Mémoires de la Société Entomologique de Belgique. 20: 33-58.
- Franco W., N. Ladino, J. H. C. Delabie, A. Dejean, J. Orivel, M. Fichaux, S. Groc, M. Leponce, and R. M. Feitosa. 2019. First checklist of the ants (Hymenoptera: Formicidae) of French Guiana. Zootaxa 4674(5): 509-543.
- Gallardo A. 1916. Las hormigas de la República Argentina. Subfamilia Dolicoderinas. Anales del Museo Nacional de Historia Natural de Buenos Aires 28: 1-130.
- Garcia M. A. The vulnerability of leaflitter ants to forest disturbances in the islands of Puerto Rico, Greater Antilles. Novitates Caribaea 13: 74-91.
- Kempf W. W. 1978. A preliminary zoogeographical analysis of a regional ant fauna in Latin America. 114. Studia Entomologica 20: 43-62.
- Kempf, W.W. 1972. Catalago abreviado das formigas da regiao Neotropical (Hym. Formicidae) Studia Entomologica 15(1-4).
- Longino J. et al. ADMAC project. Accessed on March 24th 2017 at https://sites.google.com/site/admacsite/
- Luederwaldt H. 1918. Notas myrmecologicas. Rev. Mus. Paul. 10: 29-64.
- Lutinski J. A., B. C. Lopes, and A. B. B.de Morais. 2013. Diversidade de formigas urbanas (Hymenoptera: Formicidae) de dez cidades do sul do Brasil. Biota Neotrop. 13(3): 332-342.
- Maes, J.-M. and W.P. MacKay. 1993. Catalogo de las hormigas (Hymenoptera: Formicidae) de Nicaragua. Revista Nicaraguense de Entomologia 23.
- Osorio-Perez K., M. F. Barberena-Arias, and T. M. Aide. 2007. Changes in Ant Species Richness and Composition During Plant Secondary Succession in Puerto Rico. Caribbean Journal of Science 43(2): 244-253.
- Rodrigues de Souza D., E. Stingel, L. C. de Almeida, M. A. Lazarini, C. de Bortoli Munhae, A. J. Mayhe-Nunes, O. Correa Bueno, and M. Santina de C. Morini. 2010. Ant Diversity in a Sugarcane Culture without the Use of Straw Burning in Southeast, São Paulo, Brazil. American Journal of Agricultural and Biological Sciences 5 (2): 183-188.
- Rodrigues de Souza D., E. Stingel, L. C. de Almeida, M. A. Lazarini, C. de Bortoli Munhae, O. Correa Bueno, C. R. Archangelo, and M. Santina de C. Morini. 2010. Field methods for the study of ants in sugarcane plantations in Southeastern Brazil. Sci. Agric. (Piracicaba, Braz.) 67(6): 651-657.
- Rosa da Silva R. 1999. Formigas (Hymenoptera: Formicidae) do oeste de Santa Catarina: historico das coletas e lista atualizada das especies do Estado de Santa Catarina. Biotemas 12(2): 75-100.
- Rosumek, F.B., M.A. Ulyssea, B.C. Lopes, J. Steiner. 2008. Formigas de solo e de bromélias em uma área de Mata Atlântica, Ilha de Santa Catarina, sul do Brasil: Levantamento de espécies e novos registros. Revista Biotemas 21(4):81-89.
- Santos-Junior L. C., J. M. Saraiva, R. Silvestre, and W. F. Antonialli-Junior. 2014. Evaluation of Insects that Exploit Temporary Protein Resources Emphasizing the Action of Ants (Hymenoptera, Formicidae) in a Neotropical Semi-deciduous Forest. Sociobiology 61(1): 43-51
- Shattuck S. O. 1994. Taxonomic catalog of the ant subfamilies Aneuretinae and Dolichoderinae (Hymenoptera: Formicidae). University of California Publications in Entomology 112: i-xix, 1-241.
- Silva R.R., and C. R. F. Brandao. 2014. Ecosystem-Wide Morphological Structure of Leaf-Litter Ant Communities along a Tropical Latitudinal Gradient. PLoSONE 9(3): e93049. doi:10.1371/journal.pone.0093049
- Siqueira de Castro F., A. B. Gontijo, W. Duarte da Rocha, and S. Pontes Ribeiro. 2011. As comunidades de formigas de serapilheira nas florestas semidecíduas do Parque Estadual do Rio Doce, Minas Gerais. MG.BIOTA, Belo Horizonte 3(5): 5-24.
- Smith M. R. 1937. The ants of Puerto Rico. Journal of Agriculture of the University of Puerto Rico 20: 819-875.
- Smith M. R. 1942. The relationship of ants and other organisms to certain scale insects on coffee in Puerto Rico. Journal of Agriculture of the University of Puerto Rico 26: 21-27.
- Souza D. R. de., T. T. Fernandes, J. R. de Oloveira Nascimento, S. S. Suguituru, and M. S. de C. Morini. 2012. Characterization of ant communities (Hymenoptera Formicidae) in twigs in the leaf litter of the Atlantic rainforest and Eucalyptus trees in the southeast region of Brazil. Psyche 2012(532768): 1-12
- Suguituru S. S., D. R. de Souza, C. de Bortoli Munhae, R. Pacheco, and M. S. de Castro Morini. 2011. Diversidade e riqueza de formigas (Hymenoptera: Formicidae) em remanescentes de Mata Atlântica na Bacia Hidrográfica do Alto Tietê, SP. Biota Neotrop. 13(2): 141-152.
- Suguituru S. S., M. Santina de Castro Morini, R. M. Feitosa, and R. Rosa da Silva. 2015. Formigas do Alto Tiete. Canal 6 Editora 458 pages
- Suguituru S. S., R. Rosa Silva, D. R. de Souza, C. de Bortoli Munhae, and M. Santina de Castro Morini. Ant community richness and composition across a gradient from Eucalyptus plantations to secondary Atlantic Forest. Biota Neotrop. 11(1): 369-376.
- Torres J.A. 1984. Niches and Coexistence of Ant Communities in Puerto Rico: Repeated Patterns. Biotropica 16(4): 284-295.
- Torres, Juan A. and Roy R. Snelling. 1997. Biogeography of Puerto Rican ants: a non-equilibrium case?. Biodiversity and Conservation 6:1103-1121.
- Ulyssea M. A., B. C. Lopes, A. Zillidens, J. Steiner. 2007. Ants associated to Nidularium innocentii and Aechmea lindenii (Bromeliaceae) in atlantic rainforest in southern Brazil. Biológico, São Paulo 69(2): 319-324.
- Wetterer J.K. and J.L.W. Keularts. 2008. Population explosion of the hairy crazy ant, Paratrechina pubens (Hymenoptera: Formicidae), on St. Croix, US Virgin Islands. Florida Entomologist 91(3): 423-427.
- Wetterer, J.K. and J.L.W. Keularts. 2008. Population Explosion of the Hairy Crazy Ant, Paratrechina pubens (Hymenoptera: Formicidae), on St. Croix, US Virgin Islands. The Florida Entomologist 91(3):423-427
- Wheeler W. M. 1905. The ants of the Bahamas, with a list of the known West Indian species. Bulletin of the American Museum of Natural History 21: 79-135.
- Wheeler W. M. 1908. The ants of Porto Rico and the Virgin Islands. Bulletin of the American Museum of Natural History 24: 117-158.
- Wheeler W. M. 1917. Jamaican ants collected by Prof. C. T. Brues. Bulletin of the Museum of Comparative Zoology 61: 457-471.
- Wheeler W. M. 1918. Ants collected in British Guiana by Mr. C. William Beebe. Journal of the New York Entomological Society 26: 23-28.
- Wheeler W. M. 1942. Studies of Neotropical ant-plants and their ants. Bulletin of the Museum of Comparative Zoology 90: 1-262.
- Wheeler, William Morton. 1911. Additions to the Ant-Fauna of Jamaica. Bulletin American Museum of Natural History. 30:21-29.
- Wheeler, William Morton. 1911. Ants Collected in Grenada, W.I. by Mr. C. T. Brues. Bulletin of the Museum of Comparitive Zoology at Harvard College. 54(5):166-172.
- Wild A. L. 2007. Taxonomic revision of the ant genus Linepithema (Hymenoptera: Formicidae). University of California Publications in Entomology 126: 1-151
- Wild A. L. 2009. Evolution of the Neotropical ant genus Linepithema. Systematic Entomology 34: 49-62
- Wild, A. L.. "A catalogue of the ants of Paraguay (Hymenoptera: Formicidae)." Zootaxa 1622 (2007): 1-55.