Stenamma brevicorne
| Stenamma brevicorne | |
|---|---|

| |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Arthropoda |
| Class: | Insecta |
| Order: | Hymenoptera |
| Family: | Formicidae |
| Subfamily: | Myrmicinae |
| Tribe: | Stenammini |
| Genus: | Stenamma |
| Species: | S. brevicorne |
| Binomial name | |
| Stenamma brevicorne (Mayr, 1886)
| |
| Synonyms | |
| |


From Smith (1957): Stenamma brevicorne is not only one of the most widely distributed but the oldest and perhaps best known of our North American species of Stenamma. Although apparently preferring to nest in wooded areas, the species is also found in meadows and other places. It usually nests in moderately dense to dense woods of such composition as maple, mixed oaks, oak-maple, red and white oaks, and beech-maple. Here the ants more commonly nest in the soil under stones, logs, humus, moss or other debris, as well as in the rotting wood of logs and stumps. Colonies are small, consisting of only a few dozen to a hundred or so adult individuals. The largest that has come to my attention was found at Tiffin, Ohio, on August 21, 1948, by Talbot and Headley and was composed of 105 workers, 8 alate females, and 12 males. Although no one has made a detailed study of Stenamma brevicorne, random observations indicate that this species is timid and sluggish, subterranean or hypogaeic. The workers are undoubtedly carnivorous, but may also be predaceous. Wheeler, so far as I am aware, was the first individual to advance the theory that, in some colonies at least, winged females and males overwinter in the parental nest, these sexual castes having been produced from late summer to early fall. None of the evidence I have seen disproves this, but no doubt the age of the colony has a great deal to do with whether or not sexual castes can be produced. As the colonies are not large, it is likely that there is only one mother queen per colony. This also seems to be the case in many, if not most of our own species. The overwintering sexual castes apparently start emerging from the parental colony in spring or early summer of the following year. It is believed that it is then that mating and the forming of new colonies are started. H. V. Weems, Jr., captured a copulating male and female on May 14, 1950. Since alate females have been captured at light traps and males taken while flying at dusk, it appears that both castes fly freely at night, like many other ants. Although altitudunal records are lacking for Stenamma brevicorne, it appears that this form lives in low lands or areas of only moderately high elevation.
Photo Gallery


.JPG)
{kind=link}
Identification
The worker of Stenamma brevicorne may be distinguished by its moderately large eye, which usually bears 8-10 ommatidia in its greatest diameter; the strong mesoepinoraI impression which is frequently twice as long as deep; base of epinotum usually with a transverse welt following the mesoepinorai impression; the distinct spines which are approximately one-fourth to one-third the length of the base of the epinotum; petiolar node subconical, when viewed from behind; postpetiolar node, from above, stout, as broad as, or broader than long; head, thorax, petiole and postpetiole subopaque; promesonotum largely coarsely longitudinaIly rugulose; body usually brown with the gaster light at the base and apex and darker medianly. (Smith 1957)
Distribution
CANADA, USA. From Nova Scotia, Quebec, and Ontario south to Tennessee and west to Nebraska and Minnesota.
Latitudinal Distribution Pattern
Latitudinal Range: 46.77° to 33.23583333°.
| North Temperate |
North Subtropical |
Tropical | South Subtropical |
South Temperate |
- Source: AntMaps
Distribution based on Regional Taxon Lists
Nearctic Region: United States (type locality).
Distribution based on AntMaps
Distribution based on AntWeb specimens
Check data from AntWeb
Countries Occupied
| Number of countries occupied by this species based on AntWiki Regional Taxon Lists. In general, fewer countries occupied indicates a narrower range, while more countries indicates a more widespread species. |

|
Estimated Abundance
| Relative abundance based on number of AntMaps records per species (this species within the purple bar). Fewer records (to the left) indicates a less abundant/encountered species while more records (to the right) indicates more abundant/encountered species. |
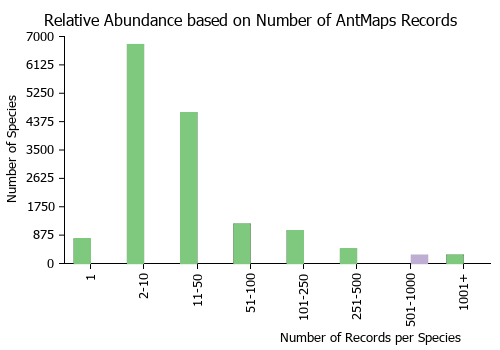
|
Biology
Regional Notes
Missouri
Talbot (1965) offers the following observations about Stenamma brevicorne from Missouri:
Stenamma brevicorne, together with Lasius brevicornis, seemed the most typical ant of the low field habitat. It was common and widespread here (23 colonies on 18 plots), but had been collected only once before on the Reserve (at swamp edge). Three other species of Stenamma (Stenamma impar, Stenamma diecki, Stenamma schmittii) present on the Reserve did not live in the field.
Stenamma brevicorne workers foraged over the ground in cool damp weather, and the finding of workers just below the surface indicates that they also foraged below ground in the crumbly topsoil. Nest entrances were simply small openings hidden among leaves and grasses, and were not found unless workers were seen entering them. An attempt was made to collect whole colonies; therefore nests were dug completely when possible. This was not always a successful process, but the general pattern of nest structure seemed clear. Usually there was a chamber within 2 inches of the surface, which might be empty during dry weather. Some nests appeared to have no chambers in the crumbly surface soil but started with a chamber 4 inches down in more solid clay. No colony had more than 4 chambers, and most had only 2. Lowest chambers ranged in depth from 3 to 13 inches, with most at 6 inches. Large colonies dug down to soil moisture, while small colonies did not. Chambers were surprisingly large and were often only partly filled with brood. Those measured ranged from 1/2 X 1/2X 1/4 in. to 1 1/2 X 1 1/2 X 1 in. with most being larger than an inch in diameter and a half inch high. The range of size of the 10 most complete population counts is recorded in Table II. Eggs, larvae and worker pupae were present from before June 23, when the first colonies were dug, through the rest of the summer, although the September 10 colony had only one worker pupa. Twelve colonies which had winged adults or winged pupae averaged 20.6 alates per colony, of which 11.2 were females and 9.4 were males. Winged pupae were found in colonies from July 9 to August 11, and adults from August 10 to September 10 when observations ceased. It is known that S. brevicorne has flights in the spring (Smith, 1957; Kannowski, 1958). It may also fly in the late fall when conditions are right.
This species generally inhabits open areas, with either a soil nest that opens underneath an object resting on the ground or within the litter horizon in debri or other objects where they can aggregate within chamber like structures.
Reproduction
Kannowski (1958) offers these detailed observations on a mating flight:
On the afternoon of May 2, 1955, I observed swarming of Stenamma brevicorne alates near "Blanchard's Pond" on the outskirts of Ann Arbor, Michigan. The pond, about an acre in size, occupied a shallow depression surrounded by a field which had not been cultivated in the last five years. Around the periphery of the pond were isolated clumps of shrubs and small trees, of which willows (Salix spp.) and hawthorns (Crataegus spp.) were the most numerous. About 150 yards to the northeast of the pond was a large oak-hickory woodland ("Eber White Woods").
I discovered the swarms about 2:45 p.m. It was a cloudy and moderately warm day (about 70 F.) with very little wind. The swarms had formed at the tops of 8 to 12 foot high hawthorns and willows at the edge of the pond. The hawthorns were in bloom. When I first sighted the swarms from a distance, I assumed that they were composed of bees attracted to the pollen of the flowers. Closer examination revealed that they were alate ants. The alates were not in contact with the trees, but always remained slightly above the tree tops. About a dozen such swarms were found at the tops of these trees. Usually, there was only one swarm per tree top, but several large trees had two swarms each.
Each swarm was composed of 15 to 50 males. These swarms were observed for an hour and a half, but no females were seen. At 4:15 p.m. it was necessary for me to leave the area, and the observations were terminated. No new swarms were seen to form during the time of observation. Each swarm hovered in approximately the same position during this time. The size varied from swarm to swarm, but each occupied an area that was roughly cuboidal in shape and measured from one to two feet on a side. Movement of an entire swarm seemed restricted to a distance of about one foot in any direction. Within each swarm the males were in continual motion without moving in any set direction.
Stenamma brevicorne colonies seldom comprise more than 200-300 individuals. Probably not more than a few dozen alates are produced each season in a single colony. Thus, the large number of males in these swarms a total of at least 250 indicates that they came from a number of colonies. I attempted to find the sources of these alates without success. I had previously found this species nesting only in wooded areas in southeastern Michigan. However, parts of the field near the pond were cursorily examined for Stenamma nests, but none was found. I next examined a small area in Eber White Woods where I had previously collected S. brevicorne. Again I found no evidence of the species. Nevertheless, I believe that these alates most likely came from colonies in Eber White Woods and/or in other nearby woods.
The occurrence of exclusively male swarms is significant. Perhaps they developed because only males took part in flights on this day. The first flights of the season of some species of ants are mostly, if not exclusively, male ones. Possibly this is also true of S. brevicorne. On the other hand, both sexes may have taken part in the flights with the males forming the swarms first and the females being attracted later in the day (The only female of S. brevicorne that I have taken in flight was collected over a lawn at the Edwin S. George Reserve, Livingston County, Michigan, about 6:15 p.m. on May 30, 1956). If the latter hypothesis has any validity, it is necessary to account for the activities of the females from the time of flight until the time they join the swarms.
The swarms were observed in an environment quite distinct from that in which nests of this species were located in this region. Evidently the primary attraction for these alates was the top of a tree, not the major environment. Perhaps swarms form at the tops of trees in any local environment. They would, however, be most noticeable where the trees were small and scattered. Additional swarms may form over the trees in the woodlands, but, because of the height of the trees and the dense foliage, they would be difficult to observe. If the alates were attracted to the tree-tops as prominences, the fact that the trees surrounding the pond were isolated may be important. In a more or less open environment any tree-top would likely be a conspicuous promontory. In a woods probably only those individual trees which towered above the general canopy level would attract the alates.
Life History Traits
- Queen number: monogynous (Rissing and Pollock, 1988; Frumhoff & Ward, 1992)
Castes
Nomenclature
The following information is derived from Barry Bolton's Online Catalogue of the Ants of the World.
- brevicorne. Aphaenogaster brevicornis Mayr, 1886d: 447 (w.q.) U.S.A. (Virginia).
- Type-material: syntype workers (number not stated), 1 syntype queen.
- Type-locality: U.S.A.: Virginia (T. Pergande).
- Type-depositories: NHMW, USNM.
- Emery, 1895c: 299 (m.); Smith, M.R. 1957b: 167 (m.); Hauschteck, 1962: 219 (k.).
- Combination in Stenamma: Emery, 1895c: 299.
- Status as species: Cresson, 1887: 260; Dalla Torre, 1893: 100; Emery, 1895c: 299; Forel, 1901e: 347; Wheeler, W.M. 1903d: 166 (in key); Wheeler, W.M. 1904e: 302; Wheeler, W.M. 1905f: 382; Wheeler, W.M. 1906b: 4; Wheeler, W.M. 1910g: 565; Wheeler, W.M. 1916m: 585; Emery, 1921f: 54; Menozzi, 1932b: 311; Cole, 1936a: 36; Wing, 1939: 162; Wesson, L.G. & Wesson, R.G. 1940: 93; Buren, 1944a: 284; Creighton, 1950a: 136; Smith, M.R. 1951a: 794; Smith, M.R. 1957b: 163 (redescription); Smith, M.R. 1958c: 116; Smith, M.R. 1967: 352; Francoeur, 1977b: 206; Smith, D.R. 1979: 1358; Allred, 1982: 506; DuBois & LaBerge, 1988: 142; Wheeler, G.C., et al. 1994: 304; Bolton, 1995b: 393; Coovert, 2005: 40; Ellison, et al. 2012: 319.
- Senior synonym of neoarcticum: Mayr, 1887: 628; Dalla Torre, 1893: 100; Emery, 1895c: 299; Creighton, 1950a: 136; Smith, M.R. 1957b: 165; Smith, M.R. 1958c: 116; Smith, D.R. 1979: 1358; Bolton, 1995b: 393.
- Distribution: Canada, U.S.A.
- neoarcticum. Stenamma neoarcticum Mayr, 1886d: 454 (w.q.m.) U.S.A. (New Hampshire, Virginia, California).
- Type-material: 2 syntype queens, 2 syntype males.
- [Note: 2 syntype workers (1 New Hampshire (T. Pergande), 1 Virginia (T. Pergande)) were excluded from the original type-series by Mayr, 1887: 628, and Emery, 1895c: 299, who identified them as specimens of S. brevicorne (Mayr), and not conspecific with the queens and males.]
- Type-locality: U.S.A.: California, Oct. (by restriction of Creighton, 1950a: 138).
- Type-depositories: NHMW, USNM.
- [Misspelled as nearcticum by Emery, 1895c: 299, Wheeler, W.M. 1903d: 166, and others.]
- Emery, 1895c: 299 (w.).
- Subspecies of westwoodii: Emery, 1895c: 299.
- Status as species: Cresson, 1887: 261; Dalla Torre, 1893: 121; Forel, 1901e: 347; Wheeler, W.M. 1903d: 166; Wheeler, W.M. 1910g: 565; Wheeler, W.M. 1917a: 519; Emery, 1921f: 54; Creighton, 1950a: 138; Smith, M.R. 1951a: 795.
- Junior synonym of brevicorne: Smith, M.R. 1957b: 165; Smith, M.R. 1958c: 116; Smith, D.R. 1979: 1358; Bolton, 1995b: 393.
- [Note: these synonymies with brevicorne all appear to relate to the excluded workers, not to the genuine syntype queens and males of neoarcticum. Snelling, R.R. 1973c: 25, identifies the queen and male syntypes of neoarcticum as synonymous with occidentale Smith, M.R., but gives occidentale priority, whereas neoarcticum should be the senior synonym.]
Type Material
- Syntype, workers, queens, Virginia (presumably near Washington, D.C.), United States, Theodore Pergande, Naturhistorisches Museum Wien, Vienna; beneath a stone (see Smith 1957).
- Syntype, 1 worker, 1 queen, Virginia (presumably near Washington, D.C.), United States, Theodore Pergande, National Museum of Natural History; beneath a stone (see Smith 1957) (USNM also has 2 workers and 4 queen apparently belonging to the original series).
Unless otherwise noted the text for the remainder of this section is reported from the publication that includes the original description.
Description
Worker
Smith (1957) - Length 2.7-3.8 mm. Funicular segments 2-6 as broad as, or broader than long; the four apical segments of the funiculus distinctly enlarged but not forming a well defined club; the length of the club exceeding the length of the remainder of the funiculus. Eye moderately large for a Stenamma, oblong, usually with approximately 8-10 ommatidia in its greatest diameter. The promesonotum, in profile, forming a rather long, uninterrupted arch in which the anterior half of the arch is distinctly more convex than the posterior half. Mesoepinotal impression usually strong and approximately twice as long as deep. Epinotum lower than the mesonotum and usually with a prominent, transverse welt near the base following the mesoepinotal impression. Base and declivity of the epinotum meeting to form a pair of short, but distinct, ruberculate spines which are approximately one-fourth to one-third the length of the base of the epinotum. Petiolar node viewed posteriorly with dorsally converging sides and truncate or feebly rounded superior border. Postpetiolar node viewed from above, stout, of variable shape, usually subspherical but occasionally subrectangular or transversely elliptical.
Frontal area and much of the clypeus smooth. Frontal region bearing fine, longitudinal and posteriorly diverging striae. Cheeks mostly longitudinally rugulose-punctate. Promesonotum usually largely, longitudinally rugulose, with the anterior border of the pronotum transversely rugulose. Dorsal surface of the epinorum with variable sculpture but quite commonly irregularly rugulose or rugulose-reticulate. Sides of thorax mostly rather coarsely, longitudinally rugulose with the exception of the lower part of the mesopleuron which is commonly reticulate-punctate. Base of gaster with distinct but short, longitudinal rugulae which are apparently never as long as the postpetiole. Petiolar and postpetiolar nodes mostly punctate or rugulose-punctate.
Head (with the exceptions of the mandibles, clypeus and frontal area), thorax, petiole and postpetiole, subopaque. Gaster smooth and shining. Propleuron occasionally somewhat shining.
Body with rather abundant, suberect to erect, grayish or light yellowish hairs of variable length. Head, and especially the gaster, usually more hairy: than the thorax. Hairs on appendages abundant but not always very closely appressed.
Head, thorax, petiole and postpetiole dark brown; antennae and legs light brown or yellowish brown. Gaster brown or blackish with the apex and the base lighter thus causing the dark area to appear as a more or less distinct, transverse band.
The head varies very noticeably in shape; with some individuals it is slender, very distinctly longer than broad whereas in other individuals the head is almost square but never quite as broad as long. The number of ommatidia in the greatest diameter of the eye is also subject to considerable variation, usually there are approximately 8-10 but one individual had only 4 or 5. Although it is common for the epinotum to bear a transverse welt near the base, the welt may be obscure or even lacking in some individuals. The postpetiolar node although usually subspherical may sometimes be transversely elliptical or even subrectangular. The sculpture on the promesonotum is usually largely longitudinally rugulose, occasionally there are individuals in which the rugulae have an arched or oblique trend or even an irregular, indescribable pattern. The color of the head, thorax, petiole and postpetiole is quite commonly reddish brown but often may be a light brown. Although the gaster is usually darker in the middle than at the base and tip there are individuals in which the base may be as dark as the median area.
Queen
Smith (1957) - Similar to the worker except as described below.
Length 3.4-4.2 mm. Ocelli small, yellowish, not noticeably protruding above the general surface of the head, the anterior ocellus located from 0.3-0.4 mm posterior to the frontal area. Greatest diameter of the eye ranging from 0.2-0.25 mm and with 14-18 ommatidia. Antennal scape failing to attain the posterior border of the head by its greatest diameter or less, funicular segments 2-7 as broad as, or broader than long; last segment of the club not as long as the combined lengths of the three preceding segments. Thorax 1.35-1.6 mm in length from the anterior border of the pronotal collar to the apices of the epinotal spines, widest anterior to the aniculations of the anterior wings where it measures 0.65-0.75 mm. Epinotal spines, in profile, variable in size and shape, usually stout and prominent and ranging from 0.10-0.15 mm in length; from above, the spines are approximately one-fourth to one-third as long as their interapical distance. Anterior wing grayish to yellowish depending upon the light, normally containing a closed cubital and a closed discoidal cell and a well developed stigma; without vein rom but with MB present, occasionally one or both of the anterior wings may have a partly or fully closed second cubital cell. MB is seldom absent but an occasional individual may lack this vein in one or both anterior wings. Upon infrequent occasions the discoidal cell may be only partly closed in one or both wings. Petiolar peduncle, in profile, often rather short and stout, the node subangular, slightly higher than long; postpetiole, in profile, approximately as high as long. Postpetiolar node, from above, convex, wider than long.
Scutellum and much of the mesonotum largely longitudinally rugulose-punctate. Pronotum, exclusive of the collar, largely transversely rugulose-punctulate. Area above the epinotal spines transversely rugulose with punctulate interspaces, the sculpturing often extending well down below the spines, occasionally however the area below the spines may be smooth or almost so.
Head, thorax, petiole, and postpetiole ranging from a light brown or reddish brown through dark brown to almost blackish brown.
Variation occurs mostly in the length, proportion, and color of the body. The variation of the wings is apparently less than other types of variation; it commonly consists in a partly or fully dosed extra cubital cell, lack of Mf3 or in the presence of a partly dosed discoidal cell.
Male
Smith (1957) - Length 3-3.5 mm. Ocelli distinct but not prominent or protruding above the general surface of the head, the anterior ocellus approximately 0.05 mm in width and located 0.2-0.25 mm posterior to the frontal area. Frontal area small but distinct, longer than broad. Frontal carinae short, 0.15-0.20 mm in length, subparallel, with scarcely any lobes. Eye large and prominent, 0.25-0.30 mm through its greatest diameter. Antennal scape (excluding the pedicel) approximately 0.28-0.37 mm in length, approximately as long as the combined lengths of the first four to six funicular segments; last funicular segment approximately as long as the combined lengths of the two preceding segments. Middle of the dorsal surface of the clypeus with a flattened area or impression that is longitudinal in direction. Mandible rather small, subtriangular. Thorax 1.1-1.5 mm in length from the anterior border of the pronotal collar to the apices of the epinotal spines, greatest width 0.6-0.75 mm just anterior to the articulations of the anterior wings. Mayrian furrows well developed, the parapsidal sutures present but best seen only in certain lights. In profile, base of epinotum depressed or flattened but the surface not horizontal, the base and declivity meeting on each side to form a broad tubercle or angle. Petiole, in profile, peduriculate, with a low not strongly convex node, which is approximately as long as the peduncle. Postpetiole, in profile, larger than the petiolar node, approximately as high as long. From above, postpetiolar node as broad as, or broader than long, usually with the appearance of being broader than long.
Head varying from rather densely and finely punctulate to reticulate-punctulate, the front with fine longitudinal striae; base of epinotum and sides of the petiole and postpetiole largely punctulate. Mesonotum and scutellum largely longitudinally rugulose-reticulate and punctate or punctulate, the area between the Mayrian furrows less heavily sculptured and therefore more shining than the mesonotum. Mesopleuron usually more weakly sculptured than the side of the epinotum which is irregularly rugulose-reticulate and punctulate. Gaster, and dorsal surface of the postpetiolar node, smooth and shining.
Pilosity light yellowish or grayish, moderately abundant, consisting of hairs of variable length which are suberect to erect.
Body brown to brownish black; mandibles, antennae, legs and apex of gaster light brown to yellowish, usually distinctly lighter than the remainder of the body.
Variations occur in the size of the male as well as in the proportions of the body. Individuals have been noted with the following abnormalities in the venation of the anterior wings: one or both wings with either a partly or else a fully developed extra cubital cell; also one or both wings lacking a dosed discoidal cell or else having only a partly dosed discoidal cell.
Karyotype
- See additional details at the Ant Chromosome Database.
 Explore: Show all Karyotype data or Search these data. See also a list of all data tables or learn how data is managed.
Explore: Show all Karyotype data or Search these data. See also a list of all data tables or learn how data is managed.
- n = 4, 2n = 8 (USA) (Hauschteck, 1962; Hauschteck, 1963).
Etymology
Morphological. Presumably brevi L., short and corn L., horn, are a reference to the spines on the propodeum.
Worker Morphology
- Explore: Show all Worker Morphology data or Search these data. See also a list of all data tables or learn how data is managed.
- Caste: monomorphic
References
- Carroll, T.M. 2011. The ants of Indiana (Hymenoptera: Formicidae). M.S. thesis, Purdue University.
- Chick, L.D., Lessard, J.-P., Dunn, R.R., Sanders, N.J. 2020. The coupled influence of thermal physiology and biotic interactions on the distribution and density of ant species along an elevational gradient. Diversity 12, 456 (doi:10.3390/d12120456).
- Davis, T. 2009. The ants of South Carolina (thesis, Clemson University).
- Emery, C. 1895d. Beiträge zur Kenntniss der nordamerikanischen Ameisenfauna. (Schluss). Zool. Jahrb. Abt. Syst. Geogr. Biol. Tiere 8: 257-360 (page 299, male described; combination in Stenamma)
- Fairweather, A.D., Lewis, J.H., Hunt, L., Smith, M.A., McAlpine, D.F. 2020. Ants (Hymenoptera: Formicidae) of Rockwood Park, New Brunswick: An assessment of species richness and habitat. Northwestern Naturalist 27(3):576–584.
- Hauschteck, E. 1962. Die Chromosomen einiger in der Schweiz vorkommender Ameisenarten. Vierteljahrsschr. Naturforsch. Ges. Zür. 107: 213-220 (page 219, karyotype described)
- Ipser, R.M., Brinkman, M.A., Gardner, W.A., Peeler, H.B. 2004. A survey of ground-dwelling ants (Hymenoptera: Formicidae) in Georgia. Florida Entomologist 87: 253-260.
- Ivanov, K. 2019. The ants of Ohio (Hymenoptera, Formicidae): an updated checklist. Journal of Hymenoptera Research 70: 65–87 (doi:10.3897@jhr.70.35207).
- Kannowski, P. B. 1958. Swarming of the ant Stenamma brevicornis (Mayr). Entomological News. 69:231-233.
- Mayr, G. 1886d. Die Formiciden der Vereinigten Staaten von Nordamerika. Verh. K-K. Zool.-Bot. Ges. Wien 36: 419-464 (page 447, worker, queen described)
- Rericha, L. 2007. Ants of Indiana. Indiana Department of Natural Resources, 51pp.
- Smith, M. R. 1957b. Revision of the genus Stenamma Westwood in America north of Mexico (Hymenoptera, Formicidae). Am. Midl. Nat. 57: 133-174 (page 167, male described; page 165, Senior synonym of neoacrticum)
- Talbot, M. 1965. Populations of ants in a low field. Insectes Sociaux. 12:19-47.
- Waters, J.S., Keough, N.W., Burt, J., Eckel, J.D., Hutchinson, T., Ewanchuk, J., Rock, M., Markert, J.A., Axen, H.J., Gregg, D. 2022. Survey of ants (Hymenoptera, Formicidae) in the city of Providence (Rhode Island, United States) and a new northern-most record for Brachyponera chinensis (Emery, 1895). Check List 18(6), 1347–1368 (doi:10.15560/18.6.1347).
- Wheeler, G. C. and J. Wheeler. 1986. The ants of Nevada. Natural History Museum of Los Angeles County, Los Angeles.
References based on Global Ant Biodiversity Informatics
- Allred D. M. 1982. Ants of Utah. The Great Basin Naturalist 42: 415-511.
- Allred, D.M. 1982. The ants of Utah. Great Basin Naturalist 42:415-511.
- Belcher A. K., M. R. Berenbaum, and A. V. Suarez. 2016. Urbana House Ants 2.0.: revisiting M. R. Smith's 1926 survey of house-infesting ants in central Illinois after 87 years. American Entomologist 62(3): 182-193.
- Buckell E. R. 1928. An annotated list of the ants of British Columbia. Proceedings of the Entomological Society of British Columbia 24: 30-34.
- Buczkowski G., and D. S. Richmond. 2012. The Effect of Urbanization on Ant Abundance and Diversity: A Temporal Examination of Factors Affecting Biodiversity. PLoS ONE 7(8): e41729. doi:10.1371/journal.pone.0041729
- Buren W. F. 1944. A list of Iowa ants. Iowa State College Journal of Science 18:277-312
- Campbell K. U., and T. O. Crist. 2017. Ant species assembly in constructed grasslands isstructured at patch and landscape levels. Insect Conservation and Diversity doi: 10.1111/icad.12215
- Canadensys Database. Dowloaded on 5th February 2014 at http://www.canadensys.net/
- Carroll T. M. 2011. The ants of Indiana (Hymenoptera: Formicidae). Master's Thesis Purdue university, 385 pages.
- Choate B., and F. A. Drummond. 2012. Ant Diversity and Distribution (Hymenoptera: Formicidae) Throughout Maine Lowbush Blueberry Fields in Hancock and Washington Counties. Environ. Entomol. 41(2): 222-232.
- Choate B., and F. A. Drummond. 2013. The influence of insecticides and vegetation in structuring Formica Mound ant communities (Hymenoptera: Formicidae) in Maine lowbush blueberry. Environ. Entomol. 41(2): 222-232.
- Clark A. T., J. J. Rykken, and B. D. Farrell. 2011. The Effects of Biogeography on Ant Diversity and Activity on the Boston Harbor Islands, Massachusetts, U.S.A. PloS One 6(11): 1-13.
- Cole A. C., Jr. 1936. An annotated list of the ants of Idaho (Hymenoptera: Formicidae). Canadian Entomologist 68: 34-39.
- Cole, A.C. 1936. An annotated list of the ants of Idaho (Hymenoptera; Formicidae). Canadian Entomologist 68(2):34-39
- Coovert G. A. 2005. The Ants of Ohio (Hymenoptera: Formicidae). Ohio Biological Survey, Inc. 15(2): 1-207.
- Coovert, G.A. 2005. The Ants of Ohio (Hymenoptera: Formicidae) Ohio Biological Survey Bulletin New Series Volume 15(2):1-196
- Davis W. T., and J. Bequaert. 1922. An annoted list of the ants of Staten Island and Long Island, N. Y. Bulletin of the Brooklyn Entomological Society 17(1): 1-25.
- Del Toro I., K. Towle, D. N. Morrison, and S. L. Pelini. 2013. Community Structure, Ecological and Behavioral Traits of Ants (Hymenoptera: Formicidae) in Massachusetts Open and Forested Habitats. Northeastern Naturalist 20: 1-12.
- Del Toro, I. 2010. PERSONAL COMMUNICATION. MUSEUM RECORDS COLLATED BY ISRAEL DEL TORO
- Drummond F. A., A. M. llison, E. Groden, and G. D. Ouellette. 2012. The ants (Formicidae). In Biodiversity of the Schoodic Peninsula: Results of the Insect and Arachnid Bioblitzes at the Schoodic District of Acadia National Park, Maine. Maine Agricultural and forest experiment station, The University of Maine, Technical Bulletin 206. 217 pages
- DuBois M. B. 1985. Distribution of ants in Kansas: subfamilies Ponerinae, Ecitoninae, and Myrmicinae (Hymenoptera: Formicidae). Sociobiology 11: 153-1353
- DuBois M. B. 1985. Distribution of ants in Kansas: subfamilies Ponerinae, Ecitoninae, and Myrmicinae (Hymenoptera: Formicidae). Sociobiology 11: 153-1354
- DuBois M. B. 1985. Distribution of ants in Kansas: subfamilies Ponerinae, Ecitoninae, and Myrmicinae (Hymenoptera: Formicidae). Sociobiology 11: 153-1355
- DuBois M. B. 1985. Distribution of ants in Kansas: subfamilies Ponerinae, Ecitoninae, and Myrmicinae (Hymenoptera: Formicidae). Sociobiology 11: 153-1356
- Dubois, M.B. and W.E. Laberge. 1988. An Annotated list of the ants of Illionois. pages 133-156 in Advances in Myrmecology, J. Trager
- Ellison A. M., E. J. Farnsworth, and N. J. Gotelli. 2002. Ant diversity in pitcher-plant bogs of Massachussetts. Northeastern Naturalist 9(3): 267-284.
- Frye J. A., T. Frye, and T. W. Suman. 2014. The ant fauna of inland sand dune communities in Worcester County, Maryland. Northeastern Naturalist, 21(3): 446-471.
- General D. M., and L. C. Thompson. 2011. New Distributional Records of Ants in Arkansas for 2009 and 2010 with Comments on Previous Records. Journal of the Arkansas Academy of Science 65: 166-168.
- Gregg R. E. 1945 (1944). The ants of the Chicago region. Annals of the Entomological Society of America 37: 447-480
- Heithaus R. E., and M. Humes. 2003. Variation in Communities of Seed-Dispersing Ants in Habitats with Different Disturbance in Knox County, Ohio. OHIO J. SCI. 103 (4): 89-97.
- Ipser R. M. 2004. Native and exotic ants (Hymenoptera: Formicidae) of Georgia: Ecological Relationships with implications for development of biologically-based management strategies. Doctor of Philosophy thesis, University of Georgia. 165 pages.
- Ivanov K. 2015. Checklist of the ants (Hymenoptera: Formicidae) of Ohio. Conference: Ohio Natural History Conference, At Columbus OH
- Ivanov, K. 2019. The ants of Ohio (Hymenoptera, Formicidae): an updated checklist. Journal of Hymenoptera Research 70: 65–87.
- Ivanov K., L. Hightower, S. T. Dash, and J. B. Keiper. 2019. 150 years in the making: first comprehensive list of the ants (Hymenoptera: Formicidae) of Virginia, USA. Zootaxa 4554 (2): 532–560.
- Kjar D. 2009. The ant community of a riparian forest in the Dyke Marsh Preserve, Fairfax County, Virginiam and a checklist of Mid-Atlantic Formicidae. Banisteria 33: 3-17.
- Kjar D., and E. M. Barrows. 2004. Arthropod community heterogeneity in a mid-Atlantic forest highly invaded by alien organisms. Banisteria 23: 26-37.
- Kjar D., and Z. Park. 2016. Increased ant (Hymenoptera: Formicidae) incidence and richness are associated with alien plant cover in a small mid-Atlantic riparian forest. Myrmecological News 22: 109-117.
- Lynch J. F. 1988. An annotated checklist and key to the species of ants (Hymenoptera: Formicidae) of the Chesapeake Bay region. The Maryland Naturalist 31: 61-106
- MacGown, J.A. and JV.G. Hill. Ants of the Great Smoky Mountains National Park (Tennessee and North Carolina).
- Mahon M. B., K. U. Campbell, and T. O. Crist. 2017. Effectiveness of Winkler litter extraction and pitfall traps in sampling ant communities and functional groups in a temperate forest. Environmental Entomology 46(3): 470–479.
- Menke S. B., E. Gaulke, A. Hamel, and N. Vachter. 2015. The effects of restoration age and prescribed burns on grassland ant community structure. Environmental Entomology http://dx.doi.org/10.1093/ee/nvv110
- Menke S. B., and N. Vachter. 2014. A comparison of the effectiveness of pitfall traps and winkler litter samples for characterization of terrestrial ant (Formicidae) communities in temperate savannas. The Great Lakes Entomologist 47(3-4): 149-165.
- Merle W. W. 1939. An Annotated List of the Ants of Maine (Hymenoptera: Formicidae). Entomological News. 50: 161-165
- Newman L. M. and R. J. Wolff. 1990. Ants of a northern Illinois Savanna and degraded savanna woodland. Procedings of the twelfth north american prairie conference. Page 71-74
- Ouellette G. D., F. A. Drummond, B. Choate and E. Groden. 2010. Ant diversity and distribution in Acadia National Park, Maine. Environmental Entomology 39: 1447-1556
- Procter W. 1938. Biological survey of the Mount Desert Region. Part VI. The insect fauna. Philadelphia: Wistar Institute of Anatomy and Biology, 496 pp.
- Smith M. R. 1957. Revision of the genus Stenamma Westwood in America north of Mexico (Hymenoptera, Formicidae). American Midland Naturalist 57: 133-174.
- Talbot M. 1976. A list of the ants (Hymenoptera: Formicidae) of the Edwin S. George Reserve, Livingston County, Michigan. Great Lakes Entomologist 8: 245-246.
- Wang C., J. Strazanac and L. Butler. 2000. Abundance, diversity and activity of ants (Hymenoptera: Formicidae) in oak-dominated mixed Appalachian forests treated with microbial pesticides. Environmental Entomology. 29: 579-586
- Wheeler G. C., J. N. Wheeler, and P. B. Kannowski. 1994. Checklist of the ants of Michigan (Hymenoptera: Formicidae). The Great Lakes Entomologist 26(4): 297-310
- Wheeler W. M. 1906. Fauna of New England. 7. List of the Formicidae. Occasional Papers of the Boston Society of Natural History 7: 1-24
- Wheeler W. M. 1906. Fauna of New England. 7. List of the Formicidae. Occasional Papers of the Boston Society of Natural History 7: 1-24.
- Wheeler, G.C., J. Wheeler and P.B. Kannowski. 1994. CHECKLIST OF THE ANTS OF MICHIGAN (HYMENOPTERA: FORMICIDAE). Great Lakes Entomologist 26:1:297-310
- Wing M. W. 1939. An annotated list of the ants of Maine (Hymenoptera: Formicidae). Entomological News 50:161-165.