Myrmica scabrinodis
| Myrmica scabrinodis | |
|---|---|

| |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Arthropoda |
| Class: | Insecta |
| Order: | Hymenoptera |
| Family: | Formicidae |
| Subfamily: | Myrmicinae |
| Tribe: | Myrmicini |
| Genus: | Myrmica |
| Species group: | scabrinodis |
| Species: | M. scabrinodis |
| Binomial name | |
| Myrmica scabrinodis Nylander, 1846
| |
| Synonyms | |
| |


A wide ranging species that occurs in a variety of habitats. In Greece it is uncommon although recorded from all mainland provinces and Ionian Islands. In northern Greece it is known from lowland and mountain habitats, in southern Greece only in mountains. In Achaia, numerous nests were observed in the border area between fir forest and pasture at an altitude 1619 m. (Borowiec & Salata, 2021)
| At a Glance | • Polygynous • Invasive |
Photo Gallery


Identification
A member of the scabrinodis complex of the scabrinodis species group. M. scabrinodis is so widespread and common that specimens are only given a superficial examination and even when workers are examined more carefully it is easy to confuse M. scabrinodis with several other scabrinodis-group species. Rarer scabrinodis-group species are often identified as M. scabrinodis (sometimes vice-versa) but most commonly workers (and sometimes queens) are confused with those of Myrmica vandeli and Myrmica specioides, and occasionally with sympatric Myrmica sabuleti populations. However, M. scabrinodis males have a much shorter scape than those of M. sabuleti and M. vandeli, and much longer erect hairs on the scape and tibiae than M. specioides, thus if males are taken with the female castes all scabrinodis-group species currently recognised, should be identifiable correctly. Thus M. scabrinodis has always been a “dust-bin” species that includes all forms that cannot be clearly discriminated by a combination of features of all three castes. Even now, we suppose that “M. scabrinodis” includes at least two (perhaps more) cryptic species that are morphologically indistinguishable but have different ecological preferences and are adapted to quite different habitats.
Collingwood (1979) - Yellow red to blackish brown according to habitat. The antennal scape is sharply angled and sinuate near the base, often with a slight lateral extension in the larger more deeply sculptured forms. The petiole has a distinctly concave anterior face which meets the truncate dorsal surface at a sharp angle. Head Index: 85.6; Frons Index: 36.8; Frontal Laminae Index: 66.5. Length: 4.0-5.0 mm.
Keys including this Species
- Key to Myrmica of West Europe and North Africa
- Key to Myrmica of East Europe, West Siberia, northern Kazakhstan, Caucasus, Asia Minor, Turkmenistan and Iran
- Key to Myrmica of species of East Siberia, Russian Far East, Mongolia, Korean Peninsula, northern China, and Japan
- Key to Myrmica of Romania
- Key to Myrmica of China
- Key to New England Myrmica
- Key to European parasitic Myrmica queens
Distribution
Throughout Europe. It has been introduced into North America where it is established on a number of the Boston Harbor Islands and possibly the nearby mainland (but occurrence there needs confirmation). It is not known from other locations in North America.
Latitudinal Distribution Pattern
Latitudinal Range: 68.138852° to 36.718333°.
| North Temperate |
North Subtropical |
Tropical | South Subtropical |
South Temperate |
- Source: AntMaps
Distribution based on Regional Taxon Lists
Nearctic Region: United States.
Palaearctic Region: Albania, Andorra, Armenia, Austria, Azerbaijan, Balearic Islands, Belarus, Belgium, Bulgaria, Channel Islands, China, Croatia, Czechia, Denmark, Estonia, Finland (type locality), France (type locality), Georgia, Germany, Greece, Hungary, Iberian Peninsula, Italy, Jersey, Kazakhstan, Kyrgyzstan, Latvia, Liechtenstein, Lithuania, Luxembourg, Montenegro, Netherlands, North Macedonia, Norway, Poland, Portugal, Republic of Korea, Republic of Moldova, Romania, Russian Federation, Serbia, Slovakia, Slovenia, Spain, Sweden, Switzerland, Türkiye, Ukraine, United Kingdom of Great Britain and Northern Ireland.
Distribution based on AntMaps
Distribution based on AntWeb specimens
Check data from AntWeb
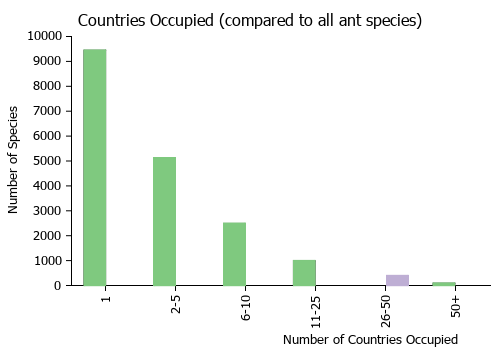
Countries Occupied
| Number of countries occupied by this species based on AntWiki Regional Taxon Lists. In general, fewer countries occupied indicates a narrower range, while more countries indicates a more widespread species. |
|
Biology
Radchenko and Elmes (2010) - Myrmica scabrinodis uses a wide variety of habitats throughout is extensive range, it is most commonly associated with grasslands but is also present in open forests and woodlands, especially in the southern part of its range. In forests it nests in the soil, often under bark and pieces of wood and sometimes in and under moss. In bogs and marshes M. scabrinodis builds nests in grass and moss tussocks and usually colonies build a quite large solarium from soil, chewed moss and other detritus, where they rear their larvae. In drier grassland and on high moorlands, nests are built under flat stones or directly into the soil, in the latter case small solaria are built in spring but these are not maintained as the season progresses. In very hot conditions nests are built in the soil with no obvious aboveground structures. In many hotter habitats that support a population of Lasius flavus mounds, nests of M. scabrinodis can be found built into the side of the mound and there is some evidence that they prey upon the L. flavus larvae, a view supported by the results of Pontin (1969) who showed that L. flavus produced more sexuals when the nests of M. scabrinodis were removed. M. scabrinodis colonies, like Myrmica rubra, have been shown to amalgamate under some conditions (Morely 1938). A cast of a M. scabrinodis nest on heathland was made by Brian and Downing (1958) and excavation behaviour was studied by Sudd (1971).

As has been pointed out in the notes above, it is hard to believe that bog-living M. scabrinodis can be the same species as sympatric populations living in very hot, dry grassland. For example, in one study (Elmes et al. 1994) we showed that on sites in both Spain and the Netherlands there appeared to be two distinct "types" of M. scabrinodis that were easily discriminated in the field. “Type I” appeared smaller and paler, lived in wetter conditions, made extensive nests in the vegetation with few if any deep chambers, were more timid being reluctant to run up ones hand if placed in the nest and attacking baits from underneath (like M. lobicornis); “type 2” appeared slightly larger and darker, lived in somewhat drier conditions, made nests with deep chambers in the soil and were more inclined to run up ones hand when placed into the nest, and sat on top of baits (more like M. ruginodis). We also noted similar ecological differences in populations from France and elsewhere in Europe (unpublished).
Subsequently we identified the “type I” specimens from Spain as Myrmica aloba, although these were somewhat smaller than is typical for that species especially compared to populations from Southern Spain and Portugal. Elsewhere, no consistent differences between the two types could be found when dead samples were compared in the laboratory; all the differences seemed to be simply behavioural. Seifert (1984) made a careful morphometric comparison in the context of M. rugulosoides (see above) and Sandor Csosz (pers. comm.) made a similar study using material from a wider range, both found no discernable morphological differences. However, some unpublished preliminary studies made by a Danish Group indicate that such populations have some genetical differences that might be sufficient to indicate cryptic species. In our view M. scabrinodis is undergoing a major speciation event at the current time largely in response to anthropogenically altered habitats (and probably also related species, such as M. aloba). It seems to us that over its range it might comprise several (perhaps four or five) cryptic species and in any region perhaps two or maybe three of these live sympatrically being separated by their behaviour and ecology. The problem is then that if an ecologist in say Britain recognises two ecological forms and another ecologist in say Hungary recognises two ecological forms there is no reason to suppose that the British pair are the same genetically as the Hungarian pair. This is a fascinating problem that cannot be resolved by normal taxonomic study, only by appropriate molecular genetical studies.
On an acid grassland site M. scabrinodis colonies average about 200 workers with 1.4 queens, 30% of nests produced sexuals and these were larger colonies with fewer queens (Elmes and Wardlaw 1982c). Like the sympatric M. sabuleti populations colony size and productivity was related to the above ground vegetation (Elmes and Wardlaw 1982a). In England, colonies on limestone grassland were the largest averaging > 4 queens and about 850 workers per nest, on acid grassland numbers were about 2 queens and 600 workers while high, cold moorland had the smallest colonies, < 0.5 queens and about 300 workers per nest; nest densities were about 1.0, 0.25 and 0.004 nests per m2 respectively (Elmes and Wardlaw 1982b). Genetical relatedness and colony structure was investigated by Seppa (1996).
The mandibular secretions of M. scabrinodis and other ants have been shown to inhibit pollen germination (Sanderson and Wright 1989) and M. scabrinodis have been shown to rob insect prey from Drosera species (Thurn 1989), which may be important for the ants when insect prey is in short supply. The general role of M. scabrinodis in moorland ant communities was studied intensively in Scotland (e.g. Brian and Brian 1951; Brian 1952a, b, 1955b, 1956b), in North Sea sand-dunes (Boomsma and De Vries 1980; Boomsma and Van Loon 1982; Boomsma and Isaaks 1982), in a mountain meadow (Woyciechowski and Miszta 1976), in steppe (Reznikova and Samoshilova 1981) and in relation to Formica ants (Vepsalanen and Savolainen 1990; Dauber and Wolters 2000), but this species has never been subjected to the same intensive study as say, M. rubra and M. ruginodis.
There have been more laboratory studies, comparable with those on the other widespread Myrmica species. For example, pheromones, glandular secretions and cuticular hydrocarbons have been identified (Cammaerts et al. 1978; Morgan et al. 1978, 1979; Elmes et al. 2002), foraging behaviour described (Dlussky et al. 1978; Cammaerts 1980b; De Vroey 1980a, b). Age related polyethism was demonstrated for M. scabrinodis (Weir 1958a, b) and the solicitation behaviour for trophollaxis was described (Lenoir 1972a, b). M. scabrinodis workers and larvae appear physiologically adapted to moderately cool conditions, at any temperature they grow at a rate intermediate between that of M. rubra and M. sabuleti (Elmes and Wardlaw 1983a). This accords to species-differences in basal respiration which vary according to the latitude at which the population lives (Nielsen et al. 1999); respiration of M. scabrinodis larvae were compared with those of Lasius flavus with which it coexists in many habitats (Peakin et al. 1985). While queens of M. scabrinodis suppress the development of large gyne-potential larvae in the same way as other Myrmica species (Elmes and Wardlaw 1983b) some gynes can develop from small over-wintered larvae and so escape queen-control (Elmes and Wardlaw 1981), which might explain why the impact of queen and worker numbers on individual size in natural colonies of M. scabrinodis can not be detected like in the cases of M. rubra and M. sabuleti (Elmes 197 4b). Also, different populations of M. scabrinodis appear more variable in the ability of the workers to oviposit (Wardlaw and Elmes 1998).
Finally, like all the other European Myrmica species nuptial flights take place from late July until the end of September (e.g. Donisthorpe 1934; Boomsma and Leusink 1981; Woyciechowski 1990a; our observations). Very often males and gynes join large mixed mating swarms but we have also observed small very local swarms composed entirely of M. scabrinodis. These local swarms often use a small bush or shrub as a swarm focus.
Collingwood (1979) - The species has variable habits, being found in a very wide range of habitats. In southern areas it is often associated with the meadow ant Lasius flavus (Fabr.) living in part of the mound nest and preying on the L. flavus workers but may be equally common in woodland, coastal sand, gravel river banks, peat bogs and moorland. Individual nests are small, situated under stones, in tree stumps or in the ground with a few hundred workers and one or a few queens. The alatas fly in August, pairing occurring in the air.
Flight Period
| X | X | X | |||||||||
| Jan | Feb | Mar | Apr | May | Jun | Jul | Aug | Sep | Oct | Nov | Dec |
Source: antkeeping.info.
- Check details at Worldwide Ant Nuptial Flights Data, AntNupTracker and AntKeeping.
 Explore: Show all Flight Month data or Search these data. See also a list of all data tables or learn how data is managed.
Explore: Show all Flight Month data or Search these data. See also a list of all data tables or learn how data is managed.
Association with Other Organisms
- Explore: Show all Associate data or Search these data. See also a list of all data tables or learn how data is managed.
Other Ants
- This species is a host for the workerless inquilines Myrmica karavajevi and Myrmica laurae.
- This species is a host for the ant Myrmica vandeli (a temporary parasite) in Europe (de la Mora et al., 2021; Jansen et al., 2010; Seifert, 2018).
- This species is a xenobiont for the ant Lasius flavus (a xenobiont) in United Kingdom (Kanizsai et al., 2013; Morley, 1945) (Foreshore. Under stone.).
Diptera
- This species is a host for the Microdon fly Microdon myrmicae (a predator) in Devon, England, U.K. (Scarparo et al., 2020).
- This species is a host for the phorid fly Pseudacteon brevicauda (a parasitoid) (Quevillon, 2018) (encounter mode primary; direct transmission; transmission outside nest).
Hemiptera
- This species is associated with the aphids Aphis asclepiadis, Aphis forbesi, Brachycaudus helichrysi, Brachycaudus persicae, Cinara brevispinosa, Cryptomyzus ribis and Drepanaphis acerifoliae (Saddiqui et al., 2019 and included references)
Hymenoptera
- This species is a host for the ichneumonid wasp Hybrizon buccatus (a parasitoid) (Quevillon, 2018) (encounter mode primary; direct transmission; transmission outside nest).
- This species is a host for the ichneumonid wasp Hybrizon buccatus (a parasitoid) (Quevillon, 2018) (encounter mode primary; direct transmission; transmission outside nest).
Lepidoptera
Myrmica scabrinodis is the only Myrmica species to be recorded as the host of all recognised European species of socially parasitic Maculinea butterflies and is one of the major hosts of Maculinea alcon and Maculinea teleius (Tartally et al., 2019; Ebsen et al., 2019).
- This species is a host for the butterfly Glaucopsyche alexis (Obregon et al. 2015).
- This species is a host for the butterfly Maculinea alcon (Tartally et al., 2019; Ebsen et al., 2019).
- This species is a host for the butterfly Maculinea teleius (Tartally et al., 2019; Ebsen et al., 2019).
Fungi
- This species is a host for the fungus Rickia wasmanniii (a pathogen) (Espadaler & Santamaria, 2012).
- This species is a host for the fungus Rickia wasmanniii (a parasite) (Quevillon, 2018) (encounter mode primary; direct transmission; transmission within nest).
Nematodes
- This species is the host for an "Mermis" (Csosz, 2012), the infected ant being treated as a distinct species under the name Sommimyrma symbiotica.
- This species is a host for the nematode Mermithidae (unspecified "Mermix") (a parasite) in Germany (Gosswald, 1930; Laciny, 2021).
Cestoda
- This species is a host for the cestode Davaineidae (Raillietina urogalli??) (a parasite) in Scotland (Muir, 1954; Laciny, 2021).
- This species is a host for the cestode Raillietina urogalli (a parasitoid) (Quevillon, 2018) (encounter mode secondary; indirect transmission; transmission outside nest).
Life History Traits
- Queen number: polygynous (Rissing and Pollock, 1988; Frumhoff & Ward, 1992)
Castes
Worker
Images from AntWeb
   
| |
| Worker. Specimen code casent0172759. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by CAS, San Francisco, CA, USA. |
      
| |
| Holotype of Myrmica martini. Worker. Specimen code antweb1008453. Photographer Roland Schultz, uploaded by California Academy of Sciences. | Owned by SMNG, Görlitz, Germany. |
Queen
Images from AntWeb
   
| |
| Queen (alate/dealate). Specimen code casent0172744. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by CAS, San Francisco, CA, USA. |
Male
Images from AntWeb
    
| |
| Male (alate). Specimen code casent0172760. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by CAS, San Francisco, CA, USA. |
Nomenclature
The following information is derived from Barry Bolton's Online Catalogue of the Ants of the World.
- scabrinodis. Myrmica scabrinodis Nylander, 1846a: 930 (w.q.m.) FINLAND. Wheeler, G.C. & Wheeler, J. 1953a: 119 (l.); Hauschteck, 1965: 325 (k.); Wasmann, 1891: 298 (gynandromorph). Subspecies of rubra: Forel, 1874: 76; Emery & Forel, 1879: 460; Emery, 1895c: 313; Forel, 1904b: 374; Wheeler, W.M. 1908g: 406. Status as species: Saunders, E. 1880: 215; Nasonov, 1889: 36; Emery, 1898c: 126; Emery, 1908a: 174; Bondroit, 1912: 351; Stitz, 1914: 71; Donisthorpe, 1915d: 125; Forel, 1915d: 29; Karavaiev, 1916: 504; Emery, 1916b: 120; Wheeler, W.M. 1917a: 504; Bondroit, 1918: 101; Santschi, 1921a: 110; Menozzi, 1922b: 325; Müller, 1923: 43; Finzi, 1926: 98; Karavaiev, 1926b: 95; Santschi, 1931b: 341; Bernard, 1967: 116; Tarbinsky, 1976: 41; Kutter, 1977c: 69; Arnol'di & Dlussky, 1978: 534; Collingwood, 1979: 55; Seifert, 1988b: 27; Atanassov & Dlussky, 1992: 95. Senior synonym of pilosiscapus: Kutter, 1977c: 69; Collingwood, 1979: 55; Seifert, 1988b: 27; Casevitz-Weulersse, 1990a: 137; of rugulosoides: Bernard, 1967: 116; Banert & Pisarski, 1972: 350; Collingwood, 1979: 55; Seifert, 1984b: 1; of ahngeri: Radchenko, 1994e: 79; of reticulata: Seifert, 1988b: 27; of scabrinodosabuleti: Radchenko & Elmes, 2010: 259; of symbiotica: Csősz, 2012: 28. Material of the nomen nudum rugosa referred here by Mayr, 1855: 411. See also: Radchenko, 2007: 29; Radchenko & Elmes, 2010: 259.
- martini. Myrmica martini Seifert, Yazdi & Schultz, 2014: 174, figs. 1-6 (w.) FRANCE.
- Junior synonym of scabrinodis: Seifert, 2024: 20.
- rugulosoides. Myrmica scabrinodis var. rugulosoides Forel, 1915d: 29 (w.) SWITZERLAND.
- Kutter, 1924: 8 (q.m.).
- Subspecies of scabrinodis: Kutter, 1924: 8; Santschi, 1931b: 342; Stitz, 1939: 93; Novak & Sadil, 1941: 79.
- Status as species: Bondroit, 1918: 102; Müller, 1923: 42; Finzi, 1926: 94; Baroni Urbani, 1971c: 30; Kutter, 1977c: 68.
- Junior synonym of scabrinodis: Bernard, 1967: 116; Banert & Pisarski, 1972: 350; Collingwood, 1979: 55; Seifert, 1984b: 1; Seifert, 1988b: 27; Seifert, 2024: 20.
- pilosiscapus. Myrmica pilosiscapus Bondroit, 1920a: 147, fig. 1 (w.q.m.) BELGIUM.
- [Also described as new by Bondroit, 1920b: 301.]
- Subspecies of scabrinodis: Finzi, 1926: 102; Santschi, 1931b: 343; Stitz, 1939: 94.
- Status as species: Sadil, 1952: 256; Arnol'di, 1970b: 1842; Baroni Urbani, 1971c: 26.
- Junior synonym of sabuleti: Bernard, 1967: 117; Boven, 1977: 120.
- Junior synonym of scabrinodis: Kutter, 1977c: 69; Collingwood, 1979: 55; Seifert, 1988b: 27; Casevitz-Weulersse, 1990a: 137; Seifert, 2024: 20.
- reticulata. Myrmica rolandi var reticulata Stärcke, 1942c: xxv (w.q.m.) SPAIN. [First available use of Myrmica scabrinodis st. rolandi var. reticulata Santschi, 1931b: 344; unavailable name.] Junior synonym of scabrinodis: Seifert, 1988b: 27.
- scabrinodosabuleti. Myrmica (Myrmica) scabrinodis var. scabrinodosabuleti Sadil, 1952: 253, figs. IV, 13-15; VII, 17-19 (w.m.) CZECHOSLOVAKIA. Junior synonym of sabuleti: Radchenko, 1994e: 80; of scabrinodis: Radchenko & Elmes, 2010: 259.
- symbiotica. Sommimyrma symbiotica Menozzi, 1925d: 25, fig. 1 (w.) ITALY. Combination in Myrmica: Bolton, 1988a: 4. Junior synonym of scabrinodis: Csősz, 2012: 28. See also: Kutter, 1973c: 256; Radchenko & Elmes, 2003a: 224; Radchenko & Elmes, 2010: 299.
Type Material
Radchenko and Elmes (2010) - We studied the type series of M. scabrinodis, preserved in the Nylander's collection (Helsinki) and designated the worker as the lectotype (see material examined). Originally, the lectotype worker was pinned through its gaster by a thin and blunt pin; during our investigation the specimen fell from this pin so we glued the lectotype to a cardboard triangle on a proper entomological pin.
Bezděčková et al. (2017) - Myrmica scabrinodis scabrinodosabuleti: Altogether six syntypes are deposited in National Museum, Prague, Czech Republic (all dry-mounted, glued on rectangular card labels):
- SYNTYPE (1 worker): ‘Myrmica / scabrinodis [hw] // 24. VIII. 39 / Borotín / BOHEMIA m. / l. V. Novak [hw] // 226 // Mus. Nat. Pragae Inv. [p] / 3404 [hw] // M. s. var. scabrinodo- [underlined] / sabuleti [underlined] Sadil (det. 1951) [hw]’.
- SYNTYPES (5 workers and one detached head, glued on one label with transversal red line and number 3483, hw): ‘Mor. Nová Říše u / Telče, 5.–6. 7. 1938/ Záleský lg. [hw] // Det. Dr. Záleský [p] M. scabr. [worker symbol] / scabr. [hw] // M. sc. var. scabrinodo- [underlined] / sabuleti [underlined] Sadil (det. 1951) [hw] // Mus. Nat. Pragae, Inv. [p] / 3038 [hw]’.
Remarks. The specimen data cited in the original description slightly differ in collecting dates from the labels of specimens preserved in National Museum, Prague, Czech Republic: SADIL (1952) gives 28-8-1939 as the collecting date for the material from Borotín and 6-7-1938 for the material from Nová Říše near Telč.
Myrmica pilosiscapus: Lectotype worker labeled ‘Mouthe Jura’, ‘nid dans Sphagnum tres humide’, ‘M.pilosiscapus type Bondr.’ [all three labels in Bondroits handwriting], ‘LECTOTYPE (upper specimen) desig. Radchenko & Elmes 2000’ [published by Radchenko & Elmes 2010]; paralectotypes: 1 worker, 2 gynes and 2 males on the same pin with lectotype; 1 paralectotype worker together with 4 males on another pin labeled ‘Mouthe Jura’, ‘Type’, ‘M. pilosiscapus type Bondr.’ [all three labels in Bondroits handwriting], ‘PARALECTOTYPE desig. Radchenko & Elmes 2000’; depository Muséum des sciences naturelles de Belgique, Bruxelles / Belgium
Myrmica scabrinodis: Lectotype worker (des. Radchenko 2007) plus three paralectotype workers on two pins labelled ‘Kuusamo’, ‘W. Nyland.’, ‘Mus. Fenn.’; depository Finnish Museum of Natural History, Helsinki / Finland.
Myrmica scabrinodis rugulosoides: Lectotype worker plus for paralectotype workers labeled ‘Typus’, ‘LECTOTYPE (bottom specimen) desig. Radchenko & Elmes 2000’ [published by Radchenko & Elmes 2010], ‘Tourbières de Roche’ [Forel’s handwriting], ‘Myrmica scabrinodis Nyl. v. rugulosoides For. type’ [Forel’s handwriting], ‘ANTWEB CASENT0907654’; depository Muséum d’histoire naturelle de Genève, Genève, Switzerland.
Myrmica martini: Holotype worker labeled "FRA: 44.1002° N, 7.2332° E St.-Martin-Vésubie-3.8NNW 1629 m, Larix-Pinus, clearing, Schultz 2002.05.15 – 126" and "Holotype Myrmica martini Seifert & al."; 8 worker para-types on three other pins and 120 worker paratypes in ethanol with identical locality labels and "Paratype Myrmica martini Seifert & al."; all material stored in Senckenberg Museum of Natural History Görlitz.
Taxonomic Notes
Sommimyrma symbiotica
Csosz (2012) - Bolton (1988) reported that the holotype might have been a developmentally abnormal specimen, avoiding any further speculations. According to the images (Figs. 1a, b) probably two or three nematode worms (confirmed by nematologists, Gábor Majoros and George Poinar) nearly completely fill in the gaster of the type individual. The worms are not completely developed; these parasites apparently did not attain the preadult stage. In general characteristics of external morphology the Myrmica symbiotica holotype specimen is reminiscent of the workers or mermithogenic phenotypes of M. scabrinodis, hence I conducted a PCA based morphometric comparison of the type with both infested and uninfected classes of M. scabrinodis. Results demonstrate that M. symbiotica holotype is nested in the cluster of infested M. scabrinodis individuals (Fig. 2). Based on these results I propose new junior synonymy with M. scabrinodis.
Cryptic Species
Ebsen et al. (2019), using molecular phylogenetic methods based on mitochondrial DNA, found two highly divergent paraphyletic lineages within M. scabrinodis consistent with earlier suggestions that cryptic species or ecotypes might well occur, either in association with habitat (Elmes et al., 1994, 1998; Radchenko & Elmes, 2010) or with latitude or longitude (Elmes & Clarke, 2005).
Unless otherwise noted the text for the remainder of this section is reported from the publication that includes the original description.
Description



Karyotype
- See additional details at the Ant Chromosome Database.
- Explore: Show all Karyotype data or Search these data. See also a list of all data tables or learn how data is managed.
- n = 22, 2n = 44 (Switzerland) (Hauschteck, 1965).
Etymology
Radchenko and Elmes (2010) - from a combination of the Latin words scabres = roughness or "scabby" and nodus = knot or lump, to describe the rough, scabby appearance of the surfaces of the petiole.
References
- Arnol'di, K. V.; Dlussky, G. M. 1978. Superfam. Formicoidea. 1. Fam. Formicidae - ants. Pp. 519-556 in: Medvedev, G. S. (ed.) Keys to the insects of the European part of the USSR. Vol. 3. Hymenoptera. Part 1. Opredeliteli Faune SSSR 119:3-584. (page 534, Status as species)
- Atanassov, N.; Dlussky, G. M. 1992. Fauna of Bulgaria. Hymenoptera, Formicidae. Fauna Bûlg. 22: 1-310 (page 95, Status as species)
- Baer, B. 2011. The copulation biology of ants (Hymenoptera: Formicidae). Myrmecological News 14: 55-68.
- Bagherian Yazdi, A., Munch, W. & Seifert, B. 2012. A first demonstration of interspecific hybridization in Myrmica ants by geometric morphometrics (Hymenoptera: Formicidae). Myrmecological News 17, 121-131.
- Banert, P.; Pisarski, B. 1972. Mrówki (Formicidae) Sudetów. Fragmenta Faunistica 18: 345-359 (page 350, Senior synonym of rugulosoides)
- Barbero, F., Bonelli, S., Thomas, J.A., Balletto, E., Schonrogge, K. 2009. Acoustical mimicry in a predatory social parasite of ants. Journal of Experimental Biology 212, 4084–4090. (doi:10.1242/JEB.032912).
- Barbero, F., Patricelli, D., Witek, M., Balletto, E., Casacci, L.P., Sala, M., Bonelli, S. 2012. Myrmica Ants and Their Butterfly Parasites with Special Focus on the Acoustic Communication. Psyche: A Journal of Entomology 2012, 1–11 (doi:10.1155/2012/725237).
- Báthori, F., Rádai, Z., Tartally, A. 2017. The effect of Rickia wasmanniii (Ascomycota, Laboulbeniales) on the aggression and boldness of Myrmica scabrinodis (Hymenoptera, Formicidae). Journal of Hymenoptera Research. 58:41–52. (doi:10.3897/jhr.58.13253)
- Bátori, Z., Vojtkó, A., Maák, I.E., Lőrinczi, G., Farkas, T., Kántor, N., Tanács, E., Kiss, P.J., Juhász, O., Módra, G., Tölgyesi, C., Erdős, L., Aguilon, D.J., Keppel, G. 2019. Karst dolines provide diverse microhabitats for different functional groups in multiple phyla. Scientific Reports 9, 7176 (doi:10.1038/s41598-019-43603-x).
- Baty, J.W., Bulgarella, M., Dobelmann, J., Felden, A., Lester, P.J. 2020. Viruses and their effects in ants (Hymenoptera: Formicidae). Myrmecological News 30: 213-228 (doi:10.25849/MYRMECOL.NEWS_030:213).
- Bereczki, J., Póliska, S., Váradi, A., Tóth, J.P. 2019. Incipient sympatric speciation via host race formation in Phengaris arion (Lepidoptera: Lycaenidae). Organisms Diversity, Evolution 20, 63–76 (doi:10.1007/s13127-019-00418-y).
- Bernadou, A., Fourcassié, V., Espadaler, X. 2013. A preliminary checklist of the ants (Hymenoptera, Formicidae) of Andorra. ZooKeys 277, 13–23 (doi:10.3897/zookeys.277.4684).
- Bernard, F. 1967a [1968]. Faune de l'Europe et du Bassin Méditerranéen. 3. Les fourmis (Hymenoptera Formicidae) d'Europe occidentale et septentrionale. Paris: Masson, 411 pp. (page 116, Status as species; Senior synonym of rugulosoides)
- Bezděčková, K., Bezděčka, P., Macek, J., Malenovský, I. 2017. Catalogue of type specimens of ants (Hymenoptera: Formicidae) deposited in Czech museums. Acta Entomologica Musei Nationalis Pragae 57(1): 295-308 (doi:10.1515/aemnp-2017-0076).
- Blatrix, R., Aubert, C., Decaëns, T., Berquier, C., Andrei-Ruiz, M.-C., Galkowski, C. 2020. Contribution of a DNA barcode to an assessment of the specificity of ant taxa (Hymenoptera: Formicidae) on Corsica. European Journal of Entomology 117: 420-429 (doi:10.14411/eje.2020.046).
- Bondroit, J. 1912. Fourmis de Hautes-Fagnes. Ann. Soc. Entomol. Belg. 56: 351-352 (page 351, Status as species)
- Bondroit, J. 1918. Les fourmis de France et de Belgique. Ann. Soc. Entomol. Fr. 87: 1-174 (page 101, Status as species)
- Boomsma, J.J., Leusink, A. 1981. Weather conditions during nuptial flights of four European ant species. Oecologia 50, 236–241. (doi:10.1007/bf00348045).
- Borowiec, L. 2014. Catalogue of ants of Europe, the Mediterranean Basin and adjacent regions (Hymenoptera: Formicidae). Genus (Wroclaw) 25(1-2): 1-340.
- Borowiec, L., Salata, S. 2021. Notes on ants (Hymenoptera: Formicidae) from Western Greece. Annals of the Upper Silesian Museum in Bytom Entomology 30: 1-23 (doi:10.5281/ZENODO.5571258).
- Borowiec, L., Salata, S. 2022. Notes on ants (Hymenoptera: Formicidae) of Thassos Island, Greece. Annals of the Upper Silesian Museum in Bytom, Entomology 31 (online 2): 1-15 (doi:10.5281/ZENODO.6123287).
- Brown, M.J.F., Bonhoeffer, S. 2003. On the evolution of claustral colony founding in ants. Evolutionary Ecology Research 5: 305–313.
- Bulter, I. 2020. Hybridization in ants. Ph.D. thesis, Rockefeller University.
- Cantone S. 2017. Winged Ants, The Male, Dichotomous key to genera of winged male ants in the World, Behavioral ecology of mating flight (self-published).
- Cantone S. 2018. Winged Ants, The queen. Dichotomous key to genera of winged female ants in the World. The Wings of Ants: morphological and systematic relationships (self-published).
- Carroll, T.M. 2011. The ants of Indiana (Hymenoptera: Formicidae). M.S. thesis, Purdue University.
- Casacci, L.P., Barbero, F., Slipinski, P., Witek, M. 2021. The inquiline ant Myrmica karavajevi uses both chemical and vibroacoustic deception mechanisms to integrate into its host colonies. Biology 10, 654 (doi:10.3390/ biology10070654)..
- Casevitz-Weulersse, J. 1990b. Étude systématique de la myrmécofaune corse (Hymenoptera, Formicidae) (Première partie). Bull. Mus. Natl. Hist. Nat. Sect. A Zool. Biol. Écol. Anim. (4) 12: 135-163 (page 137, Senior synonym of pilosiscapus)
- Collingwood, C. A. 1979. The Formicidae (Hymenoptera) of Fennoscandia and Denmark. Fauna Entomol. Scand. 8: 1-174 (page 55, Status as species; Senior synonym of pilosiscapus, rugulosoides)
- Collingwood, C.A., Prince, A. 1998. A guide to ants of Continental Portugal (Hymenoptera: Formicidae). Boletim da Sociedade Portuguesa de Entomologia. Supl nº5, pp 49.
- Csata, E., Czekes, Z., Eros, K., Nemet, E., Hughes, M., Csosz, S., Marko, B. 2013. Comprehensive survey of Romanian myrmecoparasitic fungi: new species, biology and distribution. North-western Journal of Zoology 9: 23-29.
- Csata, E., Dussutour, A. 2019. Nutrient regulation in ants (Hymenoptera: Formicidae): a review. Myrmecological News 29: 111-124 (doi:10.25849/MYRMECOL.NEWS_029:111).
- Csősz, S. 2012. Nematode infection as significant source of unjustified taxonomic descriptions in ants (Hymenoptera: Formicidae). Myrmecological News, 17, 27-31.
- Csősz, S., Báthori, F., Gallé, L., Lőrinczi, G., Maák, I., Tartally, A., Kovács, É., Somogyi, A.Á., Markó, B. 2021. The myrmecofauna (Hymenoptera: Formicidae) of Hungary: Survey of ant species with an annotated synonymic inventory. Insects 16;12(1):78 (doi:10.3390/insects12010078).
- Csosz, S., Marko, B., Galle, L. 2011. The myrmecofauna (Hymenoptera: Formicidae) of Hungary: an updated checklist. North-Western Journal of Zoology 7: 55-62.
- Czechowska, W., Radchenko, A. 1997. Myrmica hirsuta Elmes, 1978 (Hymenoptera, Formicidae) - a socially parasitic ant species new to Poland. Fragmenta Faunistica 40: 53-57.
- Czechowski, W., Radchenko, A., Czechowska, W. 2002. The ants (Hymenoptera, Formicidae) of Poland. MIZ PAS Warsaw.
- Czekes, Z., Radchenko, A.G., Csosz, S., sz-Len, A.S., Tausan, I., Benedek, K., Mark, B. 2012. The ant genus Myrmica Latreille, 1804 (Hymenoptera: Formicidae) in Romania: distribution of species and key for their identification. Entomologica Romanica 17: 29-50.
- de Bekker, C., Will, I., Das, B., Adams, R.M.M. 2018. The ants (Hymenoptera: Formicidae) and their parasites: effects of parasitic manipulations and host responses on ant behavioral ecology. Myrmecological News 28: 1-24 (doi:10.25849/myrmecol.news_028:001).
- Dekoninck, W., Ignace, D., Vankerkhoven, F., Wegnez, P. 2012. Verspreidingsatlas van de mieren van België. Bulletin de la Société royale belge d’Entomologie 148: 95-186.
- Depa, L. 2006. Weather conditions during nuptial flight of Manica rubida (LATREILLE, 1802) (Hymenoptera: Formicidae) in southern Poland. Myrmecological News 9: 27-32.
- Donisthorpe, H. 1915f. British ants, their life-history and classification. Plymouth: Brendon & Son Ltd., xv + 379 pp. (page 125, Status as species)
- Dubovikoff, D.A., Yusupov, Z.M. 2017. Family Formicidae - Ants. In Belokobylskij S. A. and A. S. Lelej: Annotated catalogue of the Hymenoptera of Russia. Proceedingss of the Zoological Institute of the Russian Academy of Sciences 6: 197-210.
- Ebsen, J.R., Boomsma, J.J. & Nash, D.R. 2019. Phylogeography and cryptic speciation in the Myrmica scabrinodis Nylander, 1846 species complex (Hymenoptera: Formicidae), and their conservation implications. Insect Conservation and Diversity 12: 467-480 (doi:10.1111/icad.12366).
- Emery, C. 1895d. Beiträge zur Kenntniss der nordamerikanischen Ameisenfauna. (Schluss). Zool. Jahrb. Abt. Syst. Geogr. Biol. Tiere 8: 257-360 (page 313, Race/subspecies of rubra)
- Emery, C. 1898c. Beiträge zur Kenntniss der palaearktischen Ameisen. Öfvers. Fin. Vetensk.-Soc. Förh. 20: 124-151 (page 126, Status as species)
- Emery, C. 1908a. Beiträge zur Monographie der Formiciden des paläarktischen Faunengebietes. Dtsch. Entomol. Z. 1908: 165-205 (page 174, Status as species)
- Emery, C. 1916a [1915]. Fauna entomologica italiana. I. Hymenoptera.-Formicidae. Bull. Soc. Entomol. Ital. 47: 79-275 (page 120, Status as species)
- Emery, C.; Forel, A. 1879. Catalogue des Formicides d'Europe. Mitt. Schweiz. Entomol. Ges. 5: 441-481 (page 460, Race/subspecies of rubra)
- Espadaler, X., Santamaria, S. 2012. Ecto- and Endoparasitic Fungi on Ants from the Holarctic Region. Psyche Article ID 168478, 10 pages (doi:10.1155/2012/168478).
- Finzi, B. 1926. Le forme europee del genere Myrmica Latr. Primo contributo. Boll. Soc. Adriat. Sci. Nat. Trieste 29: 71-119 (page 98, Status as species)
- Forel, A. 1874. Les fourmis de la Suisse. Systématique, notices anatomiques et physiologiques, architecture, distribution géographique, nouvelles expériences et observations de moeurs. Neue Denkschr. Allg. Schweiz. Ges. Gesammten Naturwiss. 26: 1-452 (page 76, Race/subspecies of rubra)
- Forel, A. 1904c [1903]. Note sur les fourmis du Musée Zoologique de l'Académie Impériale des Sciences à St. Pétersbourg. Ezheg. Zool. Muz. 8: 368-388 (page 374, Race/subspecies of rubra)
- Forel, A. 1915d. Fauna insectorum helvetiae. Hymenoptera. Formicidae. Die Ameisen der Schweiz. Mitt. Schweiz. Entomol. Ges. 12(B Beilage: 1-77 (page 29, Status as species)
- García, F., Arnal, J.M., Espadaler, X. 2008. Primeros registros de Myrmica bibikoffi Kutter, 1963 (Hymenoptera: Formicidae) en la Península Ibérica. Heteropterus Rev. Entomol. 8, 211–215.
- García, F., Cuesta-Segura, A.D., Espadaler, X. 2024. Myrmica babiensis sp. nov. (Hymenoptera: Formicidae), a new social parasite from the NW Iberian Peninsula. Annales Zoologici 74(1), 113-127 (doi:10.3161/00034541anz2024.74.1.006).
- General, D.E.M., Buenavente, P.A.C. 2021. The real deal: the ant species, Pheidole sauberi (Hymenoptera: Formicidae), first description of the queen and first record of a mermithergate in the Philippines. Philippine Journal of Systematic Biology 15(1): 1-8 (doi:10.26757/pjsb2021a1500x).
- Gippet, J.M.W., Colin, T., Grangier, J., Winkler, F., Haond, M., Dumet, A., Tragust, S., Mondy, N., Kaufmann, B. 2021. Land-cover and climate factors contribute to the prevalence of the ectoparasitic fungus Laboulbenia formicarum in its invasive ant host Lasius neglectus. Fungal Ecology 51, 101045 (doi:10.1016/j.funeco.2021.101045).
- Glaser, F. 2016. Artenspektrum, Habitatbindung und naturschutzfachliche Bedeutung von Ameisen (Hymenoptera, Formicidae) am Stutzberg (Vorarlberg, Österreich). inatura – Forschung 34: 26 S.
- Gösswald, K. 1930. Weitere Beiträge zur Verbreitung der Mermithiden bei Ameisen. Zoologischer Anzeiger 90: 13-27.
- Haelewaters, D., Boer, P., Noordijk, J. 2015. Studies of Laboulbeniales (Fungi, Ascomycota) on Myrmica ants: Rickia wasmanniii in the Netherlands. Journal of Hymenoptera Research 44, 39–47 (doi:10.3897/jhr.44.4951).
- Hauschteck, E. 1965. Halbe haploide Chromosomenzahl im Hoden von Myrmica sulcinodis Nyl. (Formicidae). Experientia (Basel) 21: 323-325 (page 325, karyotype described)
- Jansen, G., Savolainen, R. 2010. Molecular phylogeny of the ant tribe Myrmicini (Hymenoptera: Formicidae). Zoological Journal of the Linnean Society 160(3), 482–495 (doi:10.1111/j.1096-3642.2009.00604.x).
- Jansen, G., Savolainen, R., Vepsäläinen, K. 2010. Phylogeny, divergence-time estimation, biogeography and social parasite–host relationships of the Holarctic ant genus Myrmica (Hymenoptera: Formicidae). Molecular Phylogenetics and Evolution 561, 294–304 (doi:10.1016/j.ympev.2010.01.029).
- Kanizsai, O., Lőrinczi, G., Gallé, L. 2013. Nesting associations without interdependence: A preliminary review on plesiobiosis in ants. Psyche 2013, 238602 (doi:10.1155/2013/238602).
- Karavaiev, V. 1916a [1915]. Ants from Gadjatsh district of the government of Poltava and from the Province of Ferghana. Rus. Entomol. Obozr. 15: 496-507 (page 504, Status as species)
- Karavaiev, V. 1926b. Beiträge zur Ameisenfauna des Kaukasus, nebst einigen Bemerkungen über andere palaearktische Formen. Konowia 5: 93-109 (page 95, Status as species)
- Kiran, K., Karaman, C. 2020. Additions to the ant fauna of Turkey (Hymenoptera, Formicidae). Zoosystema 42(18), 285-329 (doi:10.5252/zoosystema2020v42a18).
- Kutter, H. 1977c. Hymenoptera, Formicidae. Insecta Helv. Fauna 6: 1-298 (page 69, Status as species; Senior synonym of pilosiscapus)
- Laciny, A. 2021. Among the shapeshifters: parasite-induced morphologies in ants (Hymenoptera, Formicidae) and their relevance within the EcoEvoDevo framework. EvoDevo 12, 2 (doi:10.1186/s13227-021-00173-2).
- Lapeva-Gjonova, A., Antonova, V. 2022. An updated checklist of ants (Hymenoptera, Formicidae) of Bulgaria, after 130 years of research. Biodiversity Data Journal 10, e95599 (doi:10.3897/bdj.10.e95599).
- Lapeva-Gjonova, A., Antonova, V., Ljubomirov, T. 2021. Ants (Hymenoptera, Formicidae) of Sarnena Sredna Gora Mountains (Bulgaria). Fauna of Sarnena Sredna Gora Mts, Part 2 ZooNotes, Supplement 10: 18-27.
- Lapeva-Gjonova, A., Kiran, K. 2012. Ant fauna (Hymenoptera, Formicidae) of Strandzha (Istranca) Mountain and adjacent Black Sea coast. North-Western Journal of Zoology 8(1), 72-84.
- Lyu, D.-P. 2006. Review of the genus Myrmica in Korea (Hymenoptera: Formicidae). Journal of Asia-Pacific Entomology 9: 189-202.
- Malagocka, J., Eilenberg, J., Jensen, A.B. 2019. Social immunity behaviour among ants infected by specialist and generalist fungi. Current Opinion in Insect Science 33, 99–104 (doi:10.1016/j.cois.2019.05.001).
- Masoni, A., Frizzi, F., Nieri, R., Casacci, L.P., Mazzoni, V., Turillazzi, S., Santini, G. 2021. Ants modulate stridulatory signals depending on the behavioural context. Scientific Reports 11, 5933 (doi:10.1038/s41598-021-84925-z).
- Mayr, G. 1855. Formicina austriaca. Beschreibung der bisher im österreichischen Kaiserstaate aufgefundenen Ameisen, nebst Hinzufügung jener in Deutschland, in der Schweiz und in Italien vorkommenden Arten. Verh. Zool.-Bot. Ver. Wien 5: 273-478 (page 411, Material of the nomen nudum rugosa referred here)
- Menozzi, C. 1922c. Contribution à la faune myrmécologique de l'Espagne. Bol. R. Soc. Esp. Hist. Nat. 22: 324-332 (page 325, Status as species)
- Mokadam, C. 2021. Native and non-native ant impacts on native fungi (M.A. thesis, Buffalo State University).
- Morley, D.W. 1945. Observations on some plesiobiotic colonies of ants (Hymenoptera), with notes on some other mixtobiotic colonies. Proceedings of the Royal Entomological Society of London 20, 1–4.
- Muir, D. 1954. Ants Myrmica rubra L. and M. scabrinodis Nylander as intermediate hosts of a cestode. Nature 173: 688-689 (doi:10.1038/173688b0).
- Müller, G. 1923b. Le formiche della Venezia Guilia e della Dalmazia. Boll. Soc. Adriat. Sci. Nat. Trieste 28: 11-180 (page 43, Status as species)
- Nasonov, N. V. 1889. Contribution to the natural history of the ants primarily of Russia. 1. Contribution to the ant fauna of Russia. Izv. Imp. Obshch. Lyubit. Estestvozn. Antropol. Etnogr. Imp. Mosk. Univ. 58: 1-78 (page 36, Status as species)
- Német, E., Czekes, Z., Markó, B., Rákosy, L. 2016. Host plant preference in the protected myrmecophilous Transylvanian Blue (Pseudophilotes bavius hungarica) butterfly (Lepidoptera: Lycaenidae) and its relationship with potential ant partners. Journal of Insect Conservation 20, 765–772 (doi:10.1007/S10841-016-9907-5).
- Nemet, E., Czekes, Z., Tausan, I., Marko, B. 2012. Contribution to the knowledge of the myrmecofauna of the Cefa Nature Park (North-Western Romania). Acta Scientiarum Transylvanica Biologia 20, 61-72.
- Novgorodova, T. 2021. Preventing transmission of lethal disease: Removal behaviour of Lasius fuliginosus (Hymenoptera: Formicidae) Towards Fungus Contaminated Aphids. Insects 12, 99. (doi:10.3390/insects12020099).
- Nylander, W. 1846a. Adnotationes in monographiam formicarum borealium Europae. Acta Societatis Scientiarum Fennicae. 2:875-944. (page 930, worker, queen, male described)
- Obregon, R., M. R. Shaw, J. Fernandez-Haeger, and D. Jordano. 2015. Parasitoid and ant interactions of some Iberian butterflies (Insecta: Lepidoptera). Shilap-Revista De Lepidopterologia. 43:439-454.
- Park, J., Park, J. 2021. Complete mitochondrial genome of the gate-keeper ant Colobopsis nipponica (Wheeler, W.M., 1928) (Formicidae: Hymenoptera). Mitochondrial DNA Part B 6, 86–88 (doi:10.1080/23802359.2020.1845581).
- Phillips, A.L., Attewell, P. J. 2011. An occurrence of Lasius sabularum (Bondroit, 1918) and Lasius umbratus (Nylander, 1846) within the same nest in Lincolnshire, England. BWARS Newsletter Autumn 2011:22-24.
- Radchenko, A. G. 1994h. Survey of the species of the rubra, rugosa, arnoldii, luteola and schencki groups of the genus Myrmica (Hymenoptera, Formicidae) from central and eastern Palearctic. Zool. Zh. 73(1 11: 72-80 (page 79, Senior synonym of ahngeri and georgia)
- Radchenko, A.G. & Elmes, G.W. 2010. Myrmica ants of the Old World. Fauna Mundi 3: 1-789.
- Radchenko, A.G., Fisher, B.L., Esteves, F.A., Martynova, E.V., Bazhenova, T.N., Lasarenko, S.N. 2023. Ant type specimens (Hymenoptera, Formicidae) in the collection of Volodymyr Opanasovych Karawajew. Communication 1. Dorylinae, Poneromorpha and Pseudomyrmecinae. Zootaxa, 5244(1), 1–32 (doi:10.11646/zootaxa.5244.1.1).
- Rericha, L. 2007. Ants of Indiana. Indiana Department of Natural Resources, 51pp.
- Santschi, F. 1921a. Notes sur les fourmis paléarctiques. II. Fourmis d'Asie Mineure récoltées par M. H. Gadeau de Kerville. Bol. R. Soc. Esp. Hist. Nat. 21: 110-116 (page 110, Status as species)
- Santschi, F. 1931c. Notes sur le genre Myrmica (Latreille). Rev. Suisse Zool. 38: 335-355 (page 341, Status as species)
- Satria, R. 2017. Taxonomy of the ant genus Odontomachus (Hymenoptera: Formicidae: Ponerinae) in the Indo-Chinese and Indo-Malayan subregions. Ph.D. thesis, Tokyo Metropolitan University.
- Saunders, E. 1880. Synopsis of the British Heterogyna and fossorial Hymenoptera. Trans. Entomol. Soc. Lond. 1880: 201-304 (page 215, Status as species)
- Savolainen, R., Vepsalainen, K. 2003. Sympatric speciation through intraspecific social parasitism. Proceedings of the National Academy of Sciences 100(12): 7169–7174.
- Scarparo, G., Wolton, R., Molfini, M., Pinna, L.C., di Giulio, A. 2020. Comparative morphology of myrmecophilous immature stages of European Microdon species (Diptera: Syrphidae): updated identification key and new diagnostic characters. Zootaxa 4789, 348–370 (doi:10.11646/zootaxa.4789.2.2).
- Schär, S., Vila, R., Menchetti, M. 2023. First record of the introduced ant Myrmica specioides in the eastern United States. Sociobiology 70(4), e9153 (doi:10.13102/sociobiology.v70i4.9153).
- Schifani, E., Castracani, C., Spotti, F.A., Giannetti, D., Ghizzoni, M., Gobbi, M., Lencioni, V., Pedrotti, L., Grasso, D.A., Mori, A. 2021. Social parasite ants in the Alps: a new site of the vulnerable Myrmica myrmicoxena and new uppermost altitudinal limit for M. microrubra. Sociobiology, 68(4): e7276 (doi:10.13102/sociobiology.v68i4.7276).
- Schifani, E., Csősz, S., Viviano, R., Alicata, A. 2021. Ant diversity on the largest Mediterranean islands: on the presence or absence of 28 species in Sicily (Hymenoptera, Formicidae). Natural History Sciences 8, 55–70 (doi:10.4081/nhs.2021.532).
- Schultner, E., Pulliainen, U. 2020. Brood recognition and discrimination in ants. Insectes Sociaux 67, 11–34 (doi:10.1007/s00040-019-00747-3).
- Seifert, B. 1984c. Nachweis einer im Freiland aufgetretenen Bastardierung von Leptothorax nigriceps Mayr und Leptothorax unifasciatus (Latr.) mittels einer multiplen Diskriminanzanalyse. Abh. Ber. Naturkundemus. Görlitz 58(7): 1-8 (page 1, Senior synonym of rugulosoides)
- Seifert, B. 1988b. A taxonomic revision of the Myrmica species of Europe, Asia Minor, and Caucasia (Hymenoptera, Formicidae). Abh. Ber. Naturkundemus. Görlitz 62(3): 1-75 (page 27, Status as species, Senior synonym of pilosiscapus, the unavailable name retidulata)
- Seifert, B. 2024. Pleistocene population differentiation in the ant Myrmica scabrinodis (Hymenoptera: Formicidae) – a taxonomic borderline case. Soil Organisms 96(1): 11-21 (doi:10.25674/357).
- Seifert, B., Yazdi, A. B. & Schultz, R. 2014. Myrmica martini sp.n. – a cryptic species of the Myrmica scabrinodis species complex (Hymenoptera: Formicidae) revealed by geometric morphometrics and nest-centroid clustering. Myrmecological News 19, 171-183.
- Siddiqui, J. A., Li, J., Zou, X., Bodlah, I., Huang, X. 2019. Meta-analysis of the global diversity and spatial patterns of aphid-ant mutualistic relationships. Applied Ecology and Environmental Research 17: 5471-5524 (doi:10.15666/aeer/1703_54715524).
- Snegovaya, N., Shigayev, C. 2021. A checklist of the ants (Insecta, Formicidae) of Azerbaijan Republic. Iranian Journal of Animal Biosystematics 17(2): 179-207 (doi:10.22067/ijab.2022.67343.1000).
- Sondej, I., Domisch, T. 2024. Impact of large-scale fire and habitat type on ant nest density and species abundance in Biebrza National Park, Poland. Forests 151, 123 (doi:10.3390/f15010123).
- Stevens, I., Vankerkhoven, F., Vanormeligen, P., Dekoninck, W. 2020. New observations of the social parasitic ant Myrmica karavajevi (Arnoldi, 1930) (Hymenoptera: Formicidae) in Belgium. Bulletin de la Société royale belge d’Entomologie 156: 113–121
- Stitz, H. 1914. Die Ameisen (Formicidae) Mitteleuropas, insbesondere Deutschlands. Pp. 1-111 in: Schröder, C. (ed.) Die Insekten Mitteleuropas insbesondere Deutschlands. Band II, Hymenopteren, 2. Teil. Stuttgart: Franckh'sche Verlagshandlung, 256 pp. (page 71, Status as species)
- Tarbinsky, Y.S. 1976. The ants of Kirghizia. Frunze: Ilim, 217 pp. (page 41, Status as species)
- Tartally, A., Somogyi, A.Á., Révész, T., Nash, D.R. 2020. Host ant change of a socially parasitic butterfly (Phengaris alcon) through host nest take-over. Insects 11, 556 (doi:10.3390/INSECTS11090556).
- Tausan, I., Dauber, J., Trica, M.R., Marko, B. 2017. Succession in ant communities (Hymenoptera: Formicidae) in deciduous forest clear-cuts - an Eastern European case study. European Journal of Entomology 114, 92–100 (doi:10.14411/eje.2017.013).
- Wasmann, E. 1891a [1890]. Einige neue Hermaphroditen von Myrmica scabrinodis und laevinodis. Stett. Entomol. Ztg. 51: 298-299 (page 298, gynandromorph described)
- Wegnez, P. 2017. Découverte de Myrmica lobicornis Nylander, 1846 et Lasius jensi Seifert, 1982, deux nouvelles espèces pour le Grand-Duché de Luxembourg (Hymenoptera: Formicidae). Bulletin de la Société royale belge d’Entomologie153, 46–49.
- Wheeler, G. C.; Wheeler, J. 1953a [1952]. The ant larvae of the myrmicine tribe Myrmicini. Psyche (Camb.) 59: 105-125 (page 119, larva described)
- Wheeler, W. M. 1908k. Comparative ethology of the European and North American ants. J. Psychol. Neurol. 13: 404-435 (page 406, Race/subspecies of rubra)
- Wheeler, W. M. 1917a. The mountain ants of western North America. Proc. Am. Acad. Arts Sci. 52: 457-569 (page 504, Status as species)
- Wiezik, M., Svitok, M., Wieziková, A., Dovčiak, M. 2013. Shrub encroachment alters composition and diversity of ant communities in abandoned grasslands of western Carpathians. Biodiversity and Conservation 22, 2305–2320 (doi:10.1007/s10531-013-0446-z).
References based on Global Ant Biodiversity Informatics
- Agosti, D. and C.A. Collingwood. 1987. A provisional list of the Balkan ants (Hym. Formicidae) and a key to the worker caste. I. Synonymic list. Mitteilungen der Schweizerischen Entomologischen Gesellschaft, 60: 51-62
- Aktaç, N. "Studies on the myrmecofauna of Turkey I. Ants of Siirt, Bodrum and Trabzon." Istanbul Universitesi Fen Fakultesi Mecmuasi. Seri B 41 (1977): 115-135.
- Alinvi, O., J. Bohlin and J. P. Ball. 2008. RETRACTED ARTICLE: Interspecific competition among ants in the boreal forest: Testing predictions from a linear hierarchical competition model. Insectes Sociaux 55(1):1-11.
- AntArea. Accessed on February 5th 2014 at http://antarea.fr/fourmi/
- Antarea (at www.antarea.fr on June 11th 2017)
- Antonov I. A. 2013. Ant Assemblages (Hymenoptera: Formicidae) of Cities of the Temperate Zone of Eurasia. Russian Journal of Ecology 44(6): 523526.
- Antonova V., and L. Penev. 2008. Classification of assemblages of ants in the green areas in Sofia City. Acta Zoologica Bulgarica 60(2): 103-110.
- Arnan X., M. Gracia, L. Comas, and J. Retana. 2009. Forest management conditioning ground ant community structure and composition in temperate conifer forests in the Pyrenees Mountains. Forest Ecology and Management 258(2): 5159.
- ArtDatabanken Bugs (via GBIG)
- Asociacion Iberica de Mirmecologia. 2014. List of species collected during the Taxomara 2014 Oviedo. Iberomyrmex 6: 23-24.
- Assing V. 1989. Die Ameisenfauna (Hym.: Formicidae) nordwestdeutscher Calluna-Heiden. Drosera 89: 49-62.
- Azuma M. 1938. A list of ants found in Osaka Prefecture, Japan. Entomol. World Tokyo 6: 238-243.
- Azuma M. 1938. A list of ants found in Osaka Prefecture, Japan. Entomological World. Tokyo 6:238-243.
- Azuma M. 1953. On the myrmecological fauna of Mt. Rokko, Hyogo Prefecture. Warera 2:1-7.
- Banert P, and B. Pisarski. 1972. Mrówki (Formicidae) Sudetów. Fragmenta Faunistica (Warsaw) 18: 345-359.
- Baroni Urbani C., and C. A. Collingwood. 1976. A Numerical Analysis of the Distribution of British Formicidae (Hymenoptera, Aculeata). Verhandlungen der Naturforschenden Gesellschaft in Basel 85: 51-91.
- Baroni Urbani C., and C. A. Collingwood. 1977. The zoogeography of ants (Hymenoptera, Formicidae) in Northern Europe. Acta Zoologica Fennica 152: 1-34.
- Barrett K. E. 1967. Ants in South Brittany. Entomologist's Record and Journal of Variation 79:112-116.
- Barrett K. E. J. 1968. Ants in western France. Entomologist 101: 153-155.
- Barrett K. E. J. 1968b. The distribution of ants in central southern England. Transactions of the Society for British Entomology 17: 235-250.
- Barrett K. E. J. 1970. Ants in France, 1968-69. Entomologist 103: 270-274.
- Baugnee J. Y. 2003. Camponotus piceus (Leach, 1825), fourmi nouvelle pour la faune belge decouverte dans le parc naturel Viroin-Hermeton (Hymenoptera: Formicidae). Bulletin S. R. B. E./K. B. V. E. 139: 219-225.
- Behr D., and K. Colln. 1993. Zur ameisenfauna (Hymenoptera, Formicidae) von Gonnersdorf (Kr. Daun). Dendrocopos 20: 148-160.
- Belskaya E., A. Gilev, and E. Belskii. 2017. Ant (Hymenoptera, Formicidae) diversity along a pollution gradient near the Middle Ural Copper Smelter, Russia. Environ Sci Pollut Res DOI 10.1007/s11356-017-8736-8
- Bernadou A., X. Espadaler, A. Le Goff, and V. Fourcassie. 2015. Ant community organization along elevational gradients in a temperate ecosystem. Insect. Soc. 62:5971
- Bernadou, A., G. Latil, V. Fourcassié, and X. Espadaler. "Les formigues de la Vall del Madriu-Perafita-Claror : diversitat i distribució." Hàbitats, 13 (2006): 10-21.
- Bernard F. 1967. Faune de l'Europe et du Bassin Méditerranéen. 3. Les fourmis (Hymenoptera Formicidae) d'Europe occidentale et septentrionale. Paris: Masson, 411 pp.
- Bernard F. 1975. Rapports entre fourmis et vegetation pres des Gorges du Verdon. Annales du Muséum d'Histoire Naturelle de Nice 2: 57-79.
- Bernard F. 1976. Écologie des fourmis des grès d'Annot, comparées à celles de la Provence calcaire. Annales du Muséum d'Histoire Naturelle de Nice 3: 33-54.
- Bezdeckova K., and P. Bezdecka. 2009. Nejvetsi polykalicka kolonie Formica foreli (Hymenoptera: Formicidae) y Ceské republice. Acta rerum naturalium 7: 121126.
- Blacker N. C. 1989. The ants (Hymenoptera, Formicidae) of the Gower Peninsula, West Glamorgan, South Wales. Entomologist's Record and Journal of Variation 101: 261-266.
- Blacker N. C. and C. A. Collingwood. 2002. Some significant new records of ants (Hymenoptera: Formicidae) from the Salisbury area, south Wiltshire, England, with a key to the British species of Lasius. British Journal of Entomology and Natural History 15: 25-46
- Blatrix R., C. Lebas, C. Galkowski, P. Wegnez, P. Pimenta, and D. Morichon. 2016. Vegetation cover and elevation drive diversity and composition of ant communities (Hymenoptera: Formicidae) in a Mediterranean ecosystem. – Myrmecological News 22: 119-127.
- Boer P. 2019. Species list of the Netherlands. Accessed on January 22 2019 at http://www.nlmieren.nl/websitepages/specieslist.html
- Boer P., W. Dekoninck, A. J. Van Loon, and F. Vankerkhoven. 2003. Lijst van mieren (Hymenoptera: Formicidae) van Belgie en Nederland, hun Nederlandse namen en hun voorkomen. Entomologische Berichten (Amsterdam) 63: 54-58.
- Boer P., W. Dekoninck, A. J. van Loon, and F. Vankerkhoven. 2003. Lijst van mieren (Hymenoptera: Formicidae) van Belgie en Nederland, hun Nederlandse namen en hun voorkomen. Entomologische Berichten 63(3): 54-57.
- Boer P., W. Dekoninck, A. J. van Loon, and F. Vankerkhoven. 2003. List of ants (Hymenoptera: Formicidae) of Belgium and The Netherlands, their status and Dutch vernacular names. Entomologische Berichten 63 (3): 54-58.
- Boer P., and J. Noordijk. 2004. De ruige gaststeekmier Myrmica hirsuta nieuw voor Nederland (Hymenoptera: Formicidae). Ned. Faun. Meded. 20: 25-32.
- Bonte D., W. Dekoninck, S. Provoost, E. Cosijns, and M. Hoffmann. 2003. Microgeographical distribution of ants (Hymenoptera: Formicidae) in coastal dune grassland and their relation to the soil structure and vegetation. Animal Biology 53(4): 367-377.
- Borowiec L. 2014. Catalogue of ants of Europe, the Mediterranean Basin and adjacent regions (Hymenoptera: Formicidae). Genus (Wroclaw) 25(1-2): 1-340.
- Borowiec L., and S. Salata. 2012. Ants of Greece - Checklist, comments and new faunistic data (Hymenoptera: Formicidae). Genus 23(4): 461-563.
- Borowiec L., and S. Salata. 2015. Pheidole symbioticaWasmann, 1909, an enigmatic supposed social parasite, is a nematodeinfested form of Pheidole pallidula(Nylander, 1849) (Hymenoptera: Formicidae: Myrmicinae). Sociobiology 62(2): 181-186.
- Borowiec L., and S. Salata. 2017. Ants of the Peloponnese, Greece (Hymenoptera: Formicidae). Polish Journal of Entomology 86: 193-236.
- Boven J. K. A. 1947. Liste de détermination des principales espèces de fourmis belges (Hymenoptera Formicidae). Bulletin et Annales de la Société Entomologique de Belgique 83: 163-190.
- Boven J. K. A. van. 1970. Myrmica faniensis, une nouvelle espèce parasite (Hymenoptera, Formicidae). Bull. Ann. Soc. R. Entomol. Belg. 106: 127-132.
- Bracko G. 2007. Checklist of the ants of Slovenia (Hymenoptera: Formicidae). Natura Sloveniae 9: 15-24
- Bracko G., K. Kiran, C. Karaman, S. Salata, and L. Borowiec. 2016. Survey of the ants (Hymenoptera: Formicidae) of the Greek Thrace. Biodiversity Data Journal 4: e7945. doi: 10.3897/BDJ.4.e7945
- Bracko, G. 2006. Review of the ant fauna (Hymenoptera:Formicidae) of Croatia. Acta Entomologica Slovenica 14(2): 131-156.
- Bracko, G. "Review of the ant fauna (Hymenoptera: Formicidae) of Croatia." Acta Entomologica Slovenica Vol 14 st (2006): 131-156.
- Brangham A. N. 1938. Additions to the wild fauna and flora of the Royal Botanic gardens, Kew: XVIII. Bulletin of Miscellaneous Information (Royal Botanic Gardens, Kew), 9: 390-396.
- Burrill A. C., and M. R. Smith. 1919. A key to the species of Wisconsin ants, with notes on their habits. Ohio Journal of Science 19: 279-292.
- Cagniant, H. "Note sur les peuplements de fourmis en forêt d'Algérie." Bulletin de la Société d'Histoire Naturelle de Toulouse 108 (1973): 386-390.
- Camlitepe Y., and N. Aktac. 1987. Trakya bolgesi orman karinca faunasi uzerinde arastirmalar. Türkiye I. Entomoloji Kongresi, 13-16 Ekim 1987, Ízmir
- Casevitz-Weulersse J. 1990. Etude Systematique de la Myrmecofaune Corse (Hymenoptera: Formicidae), deuxieme partie. Bull. Mus. Natn. Hist. Nat. Paris. 4eme serie 12, section A(2): 415-442.
- Casevitz-Weulersse J. 1990. Étude systématique de la myrmécofaune corse (Hymenoptera, Formicidae) (Première partie). Bull. Mus. Natl. Hist. Nat. Sect. A Zool. Biol. Écol. Anim. (4) 12: 135-163.
- Casevitz-Weulersse J., and C. Galkowski. 2009. Liste actualisee des Fourmis de France (Hymenoptera, Formicidae). Bull. Soc. Entomol. Fr. 114: 475-510.
- Casevitz-Weulersse J., and M. Prost. 1991. Fourmis de la Côte-d'Or présentes dans les collections du Muséum d'Histoire Naturelle de Dijon. Bulletin Scientifique de Bourgogne 44: 53-72.
- Castracani C., D. A. Grasso, A. Fanfani, and A. Mori. 2010. The ant fauna of Castelporziano Presidential Reserve (Rome, Italy) as a model for the analysis of ant community structure in relation to environmental variation in Mediterranean ecosystems. J Insect Conserv 14: 585594.
- Cerda X., R. Palacios, and J. Retana. 2009. Ant community structure in Citrus orchards in the Mediterranean basin: impoverishment as a consequence of habitat homogeneity. Environ. Entomol. 38(2): 317-324.
- Chen Z. L., S. Y. Zhou, and J. H. Huang. 2016. Seven species new to science and one newly recorded species of the ant genus Myrmica Latreille, 1804 from China, with proposal of a new synonym (Hymenoptera: Formicidae). ZooKeys 551: 85–128.
- Cherix D., and S. Higashi. 1979. Distribution verticale des fourmis dans le Jura vaudois et recensement prelimaire des bourdons (Hymenoptera, Formicidae et Apidae). Bull. Soc. Vaud. Sc. Nat. 356(74): 315-324.
- Clark A. T., J. J. Rykken, and B. D. Farrell. 2011. The Effects of Biogeography on Ant Diversity and Activity on the Boston Harbor Islands, Massachusetts, U.S.A. PloS One 6(11): 1-13.
- Cole A. C., Jr. 1937. An annotated list of the ants of Arizona (Hym.: Formicidae). [concl.]. Entomological News 48: 134-140.
- Colindre L. 2015. Les fourmis en Picardie: bilan 2014 (Hymenoptera/ Formicidae). Entomologiste Picard 26, 15 pages.
- Colindre L. 2017. Richess et utilite du cortege de fourmis en foret d'Ermenonville, Oise, Region Hauts-de-France. Association des Entomologistes de Picardie. 19 pages.
- Collingwood C. A. 1951. The distribution of ants in north-west Scotland. Scottish Naturalist 63: 45-49
- Collingwood C. A. 1951. The distribution of ants in north-west Scotland. Scottish Naturalist 63: 45-49.
- Collingwood C. A. 1955. Ants in S.W. Scotland. Entomologist's Record and Journal of Variation 67: 11-12.
- Collingwood C. A. 1956. Ant hunting in France. Entomologist 89: 106-108.
- Collingwood C. A. 1961. Ants in the Scottish Highlands. Scotish Naturalist 70: 12-21.
- Collingwood C. A. 1971. A synopsis of the Formicidae of north Europe. Entomologist 104: 150-176
- Collingwood C. A., and J. E. Satchell. 1956. The ants of the South Lake District. Journal of the Society for British Entomology 5: 159-164.
- Collingwood C. A., and J. Hughes. 1987. Ant species in Yorkshire, England. Naturalist (Leeds) 112: 95-101.
- Collingwood C., and A. Prince. 1998. A guide to ants of continental Portugal (Hymenoptera: Formicidae). Boletim da Sociedade Portuguesa de Entomologia. Suplemento 5: 1-49.
- Collingwood C.A. 1955. Ants in S.W. Scotland. Entomol.Rec. 67: 11-12
- Collingwood C.A. 1959. Ants in the Scottish Highlands. The Scottish Naturalist. 70: 12-21
- Collingwood C.A. 1959. Scandinavian Ants. Entomol. Rec. 71: 78-83
- Collingwood C.A. 1961. New Vice-County Records for British Ants. Entomologist. 73: 90-93
- Collingwood C.A. and Satchell J.E. 1956. The Ants of the South Lake District. Journal of the Society for British Entomology. 5: 159-164
- Collingwood, C. A. 1958. The ants of the genus Myrmica in Britain. Proc. R. Entomol. Soc. Lond. Ser. A 33: 65-75
- Collingwood, C. A. 1958b. A key to the species of ants (Hymenoptera, Formicidae) found in Britain. Trans. Soc. Br. Entomol. 13: 69-96
- Collingwood, C. A. 1964. The Identification of British Ants (Hym. Formicidae). Transactions of the Society for British Entomology. 16:93-121.
- Collingwood, C. A. 1974. A revised list of Norwegian ants (Hymenoptera: Formicidae). Norsk Entomologisk Tidsskrift 21: 31-35.
- Collingwood, C. A., and I. H. H. Yarrow. "A survey of Iberian Formicidae." EOS (Revista española de entomología) 44 (1969): 53-101.
- Collingwood, C. A. "The Formicidae (Hymenoptera) of Fennoscandia and Denmark." Fauna Entomologica Scandinavica 8 (1979): 1-174.
- Collingwood, C.A. 1958. A survey of Irish Formicidae. Proceedings of the Royal Irish Academy 59B:213-219
- Consani M., and P. Zangheri. 1952. Fauna di Romagna. Imenotteri - Formicidi. Memorie della Societa Entomologica Italiana 31: 38-48.
- Csosz S., B. Marko, K. Kiss, A. Tartally, and L. Galle. 2002. The ant fauna of the Ferto-Hansag National Park (Hymenoptera: Formicidae). In: Mahunka, S. (Ed.): The fauna of the Fert?-Hanság National Park. Hungarian Natural History Museum, Budapest, pp. 617-629.
- Csősz S., B. Markó, and L. Gallé. 2001. Ants (Hymenoptera: Formicidae) of Stana Valley (Romania): Evaluation of the effectiveness of a myrmecological survey. Entomologica Romanica 6 : 121-126.
- Csősz S., B. Markó, and L. Gallé. 2011. The myrmecofauna (Hymenoptera: Formicidae) of Hungary: an updated checklist. North-Western Journal of Zoology 7: 55-62.
- Cuní, M. "Fauna entomológica de la villa de Calella." Anales de la Sociedad española de Historia Natural (1897): 281.
- Czechowski W., A. Radchenko, W. Czechowska and K. Vepsäläinen. 2012. The ants of Poland with reference to the myrmecofauna of Europe. Fauna Poloniae 4. Warsaw: Natura Optima Dux Foundation, 1-496 pp
- Czekes Z., Radchenko, A. G., Csősz, S. Szász-Len, A., Tăuşan, I., Benedek, K., and Markó, B. 2013. The genus Myrmica Latreille, 1804 (Hymenoptera: Formicidae) in Romania: distribution of species and key for their identification. Entomologica Romanica 17: 29-50.
- Dauber J., and D. Simmering. 2006. Ant assemblages in successional stages of Scotch Broom stands (Hymenoptera: Formicidae; Spermatophyta). Myrmecologische Nachrichten 9: 55-64.
- Dauber, J., J. Bengtsson and L. Lenoir. 2006. Evaluating Effects of Habitat Loss and Land-Use Continuity on Ant Species Richness in Seminatural Grassland Remnants. Conservation Biology 20(4):1150-1160
- Dekoninck W., F. Hendrickx, M. Dethier, and J. P. Maelfait. 2010. Forest Succession Endangers the Special Ant Fauna of Abandoned Quarries along the River Meuse (Wallonia, Belgium). Restoration Ecology 18(5): 681690.
- Dekoninck W., H. De Koninck, J. Y. Baugnee, and J. P. Maelfait. 2007. Ant biodiversity conservation in Belgian calcareous grasslands: active management is vital. Belg. J. Zool. 137 (2): 137-146.
- Dekoninck W., K. Desender, and P. Grootaert. 2008. Establishment of ant communities in forests growing on former agricultural fields: Colonisation and 25 years of management are not enough (Hymenoptera: Formicidae). Eur. J. Entomol. 105: 681689.
- Della Santa E. 1995. Fourmis de Provence. Faune Provence 16: 5-37.
- Della Santa E. 2000. L'identification des espèces du genre Myrmica Latreille (Formicidae) de Suisse; essai de présentation synoptique. Bull. Romand Entomol. 18: 169-187.
- Dewes E. 2005. Ameisenerfassung im Waldschutzgebiet Steinbachtal/Netzbachtal. Abh. Delattinia 31: 89-118.
- Donisthorpe H. 1914. Myrmecophilous notes for 1913. Entomologist's Record and Journal of Variation 26: 37-45.
- Donisthorpe, H. "A first instalment of the ants of Turkey." Annals and Magazine of Natural History (12)3 (1950): 1057-1067.
- Du Merle P. 1978. Les peuplements de fourmis et les peuplements d'acridiens du Mont Ventoux II. - Les peuplements de fourmis. Terre Vie 32(1): 161-218.
- Dubovikoff D. A., and Z. M. Yusupov. 2018. Family Formicidae - Ants. In Belokobylskij S. A. and A. S. Lelej: Annotated catalogue of the Hymenoptera of Russia. Proceedingss of the Zoological Institute of the Russian Academy of Sciences 6: 197-210.
- Ebsen J. R., J. J. Boomsma, and D. R. Nash. 2019. Phylogeography and cryptic speciation in the Myrmica scabrinodis Nylander, 1846 species complex (Hymenoptera: Formicidae), and their conservation implications. Insect Conservation and Diversity doi: 10.1111/icad.12366
- Elmes, G.W., T. Akino, J.A. Thomas, R.T. Clarke and J.J. Knapp. 2002. Interspecific differences in cuticular hydrocarbon profiles of Myrmica ants are sufficiently consistent to explain host specificity by Maculinea (large blue) butterflies. Oecologia 130:525-535
- Else G., B. Bolton, and G. Broad. 2016. Checklist of British and Irish Hymenoptera - aculeates (Apoidea, Chrysidoidea and Vespoidea). Biodiversity Data Journal 4: e8050. doi: 10.3897/BDJ.4.e8050
- Emery C. 1869. Enumerazione dei formicidi che rinvengonsi nei contorni di Napoli con descrizioni di specie nuove o meno conosciute. Ann. Accad. Aspir. Nat. Secunda Era 2: 1-26.
- Emery C. 1916. Fauna entomologica italiana. I. Hymenoptera.-Formicidae. Bullettino della Società Entomologica Italiana 47: 79-275.
- Emery, C. "Beiträge zur Kenntniss der palaearktischen Ameisen." Öfversigt af Finska Vetenskaps-Societetens Förhandlingar (Helsinki) 20 (1898): 124-151.
- Emery, C. "Catalogo delle formiche esistenti nelle collezioni del Museo Civico di Genova. Parte seconda. Formiche dell'Europa e delle regioni limitrofe in Africa e in Asia." Annali del Museo Civico di Storia Naturale 12 (1878): 43-59.
- Entomological Society of Latvia. 2003. http://leb.daba.lv/Formicidae.htm (Accessed on December 1st 2013).
- Espadaler X., X. Roig, K. Gómez, and F. García. 2011. Formigues de les Planes de Son i mata de València (Hymenoptera, Formicidae) Treballs de la Institució Catalana d'Història Natural 16: 609-627.
- Espadaler, X. "Contribución al conocimiento de los formícidos (Hymenoptera, Formicidae) del Pirineo catalán." Tesis Universida (1979): 285 pp.
- Espadaler, X. "Formicidos de las sierras de Cazorla, del Pozo y Segura (Jaén, España)." Ecología 11 (1997): 489-499.
- Espadaler, X. "Lasius neglectus Van Loon, Boosma & Andrásfalvy, 1990 a potential pest ant in Spain." Orsis 14 (1999): 43-46.
- Finzi B. 1926. Le forme europee del genere Myrmica Latr. Primo contributo. Bollettino della Società Adriatica di Scienze Naturali in Trieste. 29: 71-119.
- Forel A. 1890. Fourmis de Tunisie et de l'Algérie orientale. Annales de la Société Entomologique de Belgique 34: lxi-lxxvi.
- Forel A. 1892. Die Ameisenfauna Bulgariens. (Nebst biologischen Beobachtungen.). 305-318.
- Forel A. 1904. Note sur les fourmis du Musée Zoologique de l'Académie Impériale des Sciences à St. Pétersbourg. Ezheg. Zool. Muz. 8: 368-388.
- Forel, A. "Ameisen aus den Sporaden, den Cykladen und Griechenland, gesammelt 1887 von Herrn von Oertzen." Berliner Entomologische Zeitschrift 32 (1889): 255-265.
- Forel, A. "Fourmis de Tunisie et de l'Algerie orientale recoltees et decrites par Auguste Forel." Annales de la Société Entomologique de Belgique 34 (1890): lxi-lxxvi.
- Forel, A. "Südpalaearctische Ameisen." Mitteilungen der Schweizerischen Entomologischen Gessellschaft 9 (1895): 227-234.
- Formidabel Database
- Fowles, A.P. 1996. A provisional checklist of the invertebrates recorded from Wales. 2. Aculeate wasps, bees and ants (Hymenoptera: Aculeata). Countryside Council for Wales
- Francois J. 1958. Contribution a l'etude ecologique des Formicides (Insectes, Hymenopteres) de la region Dijonnaise. Travaux du laboratoire de Zoologie et de la Station Aquicole Grimaldi de la Faculte des Sciences de Dijon 25, 39 pages.
- GRETIA. 2017. Bilan annuel de l'enquete sur la repartition des fourmis armoricaines. 23 pages.
- Gadeau de Kerville H. 1922. Materiaux pour la Faune des Hymenopteres de la Normandie. Bull. Soc. Amis Sc. Nat. Rouen 1916-1921, 1922: 217-225.
- Gaige F. M. 1925. Results of the Mershon expedition to the Charity Islandsm Lake Huron; the Formicidae of Charity island. Occasional Papers of the Museum of Zoology 5: 1-29.
- Galkowski C. 2010. Découverte de Myrmica vandeli Bondroit, 1920 dans les Pyrénées Orientales (Hymenoptera, Formicidae). Revue de l'Association roussillonnaise d'Entomologie 19 (1) : 11-12.
- Galkowski C., and P Wegnez. 2010. Myrmica constricta Karavaiev 1934, nouvelle espece pour la France (Hymenoptera, Formicidae). Bull. Soc. Ent. Mulhouse 66(3): 41-45.
- Galle L. 1972. Study of ant-populations in various grassland ecosystems. Acta Biologica Szeged 18(1-4): 159-164.
- Galle L. 1981. The Formicoid fauna of the Hortobagy. Pp. 307-311 in: Mahunka, S. (ed.) 1981. The fauna of the Hortobágy National Park. Budapest: Akadémiai Kiadó, 415 pp.
- Galle L. 1993. Data to the ant fauna of the Bukk (Hymenoptera: Formicidae). Natural history of the national parks of Hungary 7: 445-448.
- Gallé L., B. Markó, K. Kiss, E. Kovács, H. Dürgő, K. Kőváry, and S. Csősz. 2005. Ant fauna of Tisza river basin (Hymenoptera: Formicidae). In: Gallé, L. (szerk.): Vegetation and Fauna of Tisza River Basin I. Tiscia Monograph Series 7; Szeged, pp. 149-197.
- Garcia Garcia F., and A. D. Cuesta-Esgura. 2017. First catalogue of the ants of Burgos province, Spain (Hymenoptera: Formicidae). Boletín de la Sociedad Entomológica Aragonesa 60: 245–258.
- Gaspar C. 1966. Etude myrmecologique des tourbieres dans les Hautes-Fagnes en Belgique (Hymenoptera, Formicidae). Rev. Ecol. Biol. Sol. 3(2): 301-312.
- Gaspar C. 1966. Etude myrmecologique des tourbieres dans les Huates-Fagnes en Belgique (Hymenoptera, Formicidae). Rev. Ecol. Biol. Sol. 3(2): 301-312.
- Gaspar C., and C. Thirion. 1978. Modification des populations d'Hymenopteres sociaux dans les milieux anthropogenes. Memorabilia Zoologica 29: 61-77.
- Gaspare Charles. 1965. Étude myrmécologique d'une région naturelle de Belgique: la Famenne Survey des Fourmis de la Région (Hymenoptera, Formicidae). Institut agronomique de l'Etat a' Gembloux. 32(4): 427-434.
- Gibb H., and T. Johansson. 2011. Field tests of interspecific competition in ant assemblages: revisiting the dominant red wood ants. Journal of Animal Ecology 80: 548-557.
- Glaser F. 2009. Die Ameisen des Fürstentums Liechtenstein. (Hymenoptera, Formicidae). Amtlicher Lehrmittelverlag, Vaduz, 2009 (Naturkundliche Forschung im Fürstentum Liechtenstein; Bd. 26).
- Glaser F., A. Freitag, and H. Martz. 2012. Ants (Hymenoptera: Formicidae) in the Münstertal (Val Müstair) a hot spot of regional species richness between Italy and Switzerland. Gredleriana 12: 273 - 284.
- Glaser F., T. Kopf, and K. H. Steiberger. 2003. Ameisen (Hymenoptera: Formicidae) im Frastanzer Ried und den Illauen (Vorarlberg, Österreich) Artenspektrum, Gefährdung und Schutzempfehlungen. Vorarlberger Naturschau 13: 287-310.
- Gomez C., D. Casellas, J. Oliveras, and J. M. Bas. 2003. Structure of ground-foraging ant assemblages in relation to land-use change in the northwestern Mediterranean region. Biodiversity and Conservation 12: 21352146.
- Gouraud C. 2015. Bilan de l’année 2014 : Atlas des fourmis de Loire-Atlantique (Hymenoptera, Formicidae). Atlas des Formicidae de Loire-Atlantique, compte rendu de la première année d’étude (2014)
- Grandi G. 1935. Contributi alla conoscenza degli Imenotteri Aculeati. XV. Boll. R. Ist. Entomol. Univ. Studi Bologna 8: 27-121.
- Gratiashvili N., Barjadze S. 2008. Checklist of the ants (Formicidae Latreille, 1809) of Georgia. Proceedings of the Institute of Zoology (Tbilisi) 23: 130-146.
- Gribodo, G., Emery C. "Ordo Hymenoptera." Pp. 81-85 in: Cavanna, G. Parte II.- Catalogo degli animali raccolti al Vulture, al Pollino ed in altri luoghi dell'Italia meridionale e centrale Bull. Soc. Entomol. Ital. 14 (1882): 31-87.
- Grzes I. M. 2009. Ant species richness and evenness increase along a metal pollution gradient in the Boles?aw zinc smelter area. Pedobiologia 53: 65-73.
- Guénard B., and R. R. Dunn. 2012. A checklist of the ants of China. Zootaxa 3558: 1-77.
- Haguet G., M. Chevrier, and E. Brunel. 2002. Les invertebres de la dune de Bon Abri, premier inventaire. Groupe d'Etude des Invertebres Armoricains, 37 pages.
- Holec M., and J. Frouz. 2005. Ant (Hymenoptera: Formicidae) communities in reclaimed and unreclaimed brown coal mining spoil dumps in the Czech Republic. Pedobiologia 49: 345-357.
- Holgersen H. 1942. Ants of northern Norway (Hym., Form.). Tromso Mus. Årsh. 63(2): 1-34.
- Holgersen H. 1943. Ant studies in Rogaland (south-western Norway). Avhandlingar utgitt av det Norske Videnskaps-Akademi i Oslo. I. Matematisk-Naturvidenskapelig Klasse 1943(7): 1-75.
- Hågvar S. 2005. Altitudinal zonation of ants (Formicidae) in a steep fjord landscape in Sogndal, Western Norway. Norw. J. Entomol. 52: 3-12.
- Jakubzik A., H. Kinkler, and K. Colln. 2010. Aculeate Hymenoptera from a Humid Biotope in Leverkusen-Steinbüchel. Decheniana (Bonn) 163: 145158.
- Jansen G., R. Savolainen, K. Vespalainen. 2010. Phylogeny, divergence-time estimation, biogeography and social parasite–host relationships of the Holarctic ant genusMyrmica(Hymenoptera: Formicidae). Molecular Phylogenetics and Evolution 56: 294-304.
- Jeffery H. G. 1931. The Formicidae (or ants) of the Isle of Wight. Proceedings of the Isle of Wight Natural History and Archaeological Society 2: 125-128.
- Karaman M. G. 2011. A catalogue of the ants (Hymenoptera, Formicidae) of Montenegro. Podgorica: Catalogues 3, Volume 2, Montenegrin Academy of Sciences and Arts, 140 pp.
- Karaman M. G., and G. S. Karaman. 2005. New data on myrmecofauna (Hymenoptera, Formicidae) from Montenegro (Serbia & Montenegro). The Montenegrin Academy of Sciences and Arts. Glasnik of the Section of Natural Sciences 16: 51-68.
- Karaman M. G., and G. S. Karaman. 2007. Contribution to the Knowledge of the ants (Hymenoptera, Formicidae) from Special nature reserve Zasavica , Serbia. Zbornik “Naucno-strucni skup Zasavica 2007, Sremska Mitrovica, 67-75.
- Karavaiev V. 1926. Beiträge zur Ameisenfauna des Kaukasus, nebst einigen Bemerkungen über andere palaearktische Formen. Konowia 5: 93-109.
- Kiran K., and C. Karaman. 2012. First annotated checklist of the ant fauna of Turkey (Hymenoptera: Formicidae). Zootaxa 3548: 1-38.
- Kiss K., and K. Fetyko. 2008. Notes about the ant fauna (Hymenoptera: Formicidae) of the Tur valley. In. Sike T., Márk-Nagy J. (eds), The flora and fauna of the Tur river natural reserve, Biharean Biologist, II, Oradea, pp.71-76.
- Kofler A. 1995. Nachtrag zur Ameisenfauna Osttirols (Tirol, Österreich) (Hymenoptera: Formicidae). Myrmecologische Nachrichten 1: 14-25.
- Korlevic, A. "Prilozi fauni hrvatskih opnokrilaca." Glasn. Hrv. Narav. Dr. 5 (1890): 189-250.
- Kozisek T. 1987. Ants (Formicoidea) of the Abrod State Nature Reserve. Ochrana Prírody 8: 205-208.
- Krausse, A. H. "Ueber sardische Ameisen." Archiv für Naturgeschichte, Berlin A 78(7) (1912): 162-166.
- Kvamme T. 1982. Atlas of the Formicidae of Norway (Hymenoptera: Aculeata). Insecta Norvegiae 2: 1-56.
- Kvamme T., and A. Wetas. 2010. Revidert liste over norske maur Inkludert dialektiske navn og forslag til nye norske navn og forslag til norske navn. Norsk institutt for skog og landskap, Ås. 127 pp
- Laeger T., and R. Schultz. 2005. Ameisen (Hymenoptera: Formicidae) als Beifänge in Bodenfallen wie genau spiegeln sie reale Abundanzverhältnisse wider? Myrmecologische Nachrichten 7: 17-24.
- Lapeva-Gjonova A., and S. Santamaria. 2011. First records of Laboulbeniales (Ascomycota) on ants (Hymenoptera: Formicidae) in Bulgaria. ZooNotes 22: 1-6.
- Lapeva-Gjonova, L., V. Antonova, A. G. Radchenko, and M. Atanasova. "Catalogue of the ants (Hymenoptera, Formicidae) of Bulgaria." ZooKeys 62 (2010): 1-124.
- Le Moli F., and A. Zaccone. 1995. Ricerche sulla mirmecofauna del Cansiglio (Prealpi Carniche). Soc. Ven. Sc. Nat. 20: 33-52.
- Lebas C., C. Galkowski, P. Wegnez, X. Espadaler, and R. Blatrix. 2015. The exceptional diversity of ants on mount Coronat (Pyrénées-Orientales), and Temnothorax gredosi(Hymenoptera, Formicidae) new to France. R.A.R.E., T. XXIV (1): 24 33
- Lehouck V., D. Bonte, W. Dekoninck, and J. P. Maelfait. 2004. The distribution of ant nests (Hymenoptera, Formicidae) in coastal grey dunes of Flanders (Belgium) and their relationship to myrmecochorous plants. Belg. J. Zool. 134 (2/1) : 89-96.
- Lillig M., and E. Dewes. 2015. The former Siegfried Line as habitats for ants (Hymenoptera: Formicidae). Abh. Delattinia 37: 117 - 136
- Livory A. 2003. Les fourmis de la Manche. L'Argiope 39: 25-49.
- Livory A. Un Nouvel Inventaire: les Fourmis. Argiope 29: 27-34.
- Lomnicki J. 1928. Spis mrówek Lwowa i okolicy. Ksiegi Pamiatkowej (Lecia Gimn. IV Jana Dlugosza Lwowie) 50: 1-10.
- Lorinczi G. 2011. Lasius (Chthonolasius) nitidigaster Seifert, 1996 -a new ant species (Hymenoptera: Formicidae) for the Hungarian fauna. Natura Somogyiensis 19: 223-228.
- Maavara V. 1953. Ants of Estonian SSR. ABIKS loodusevaatlejale 10: 1-44.
- Majzlan O., and P. Devan. 2009. Selected insect groups (Hymenoptera, Neuroptera, Mecoptera, Raphidioptera) of the Rokoš Massif (Strážovské vrchy Mts.). Rosalia (Nitra), 20, p. 63–70.
- Malozemova L. A. 1972. Ants of steppe forests, their distribution by habitats, and perspectives of their utilization for protection of forests (north Kazakhstan). [In Russian.]. Zoologicheskii Zhurnal 51: 57-68.
- Marikovsky P. I. 1979. Ants of the Semireche Desert. [In Russian.]. Alma Ata: Nauka, 263 pp.
- Marko B. 2008. Ants (Hymenoptera: Formicidae) of the Sand Dunes of Foieni Protected Area and Its Surroundings (Satu Mare County, Romania), and a New Species for the Romanian Fauna. Acta Scientiarum Transylvanica 16(3): 87-99.
- Marko B., K. Kiss, and L. Galle. 2004. Mosaic structure of ant communities (Hymenoptera: Formicidae) in Eastern Carpathian marshes: regional versus local scales. Acta Zoologica Academiae Scientiarum Hungaricae 50(2): 7795.
- Markó B., B. Sipos, S. Csősz, K. Kiss, I. Boros, and L. Gallé. 2006. A comprehensive list of the ants of Romania (Hymenoptera: Formicidae). Myrmecologische Nachrichten 9: 65-76.
- Menozzi C. 1918. Primo contributo alla conoscenza della fauna mirmecologica del Modenese. Atti della Società dei Naturalisti e Matematici di Modena. (5)4: 81-88.
- Nemet E., Z. Czekes, I. Tausan, and B. Marko. 2012. Contribution to the knowledge of the myrmecofauna of the Cefa Nature Park (North-Western Romania). Acta Scientiarum Transylvanica 20(1): 61-72.
- Neumeyer R., and B. Seifert. 2005. Commented check list of free living ant (Hymenoptera: Formicidae) species of Switzerland. Bulletin de la Societe Entomologique Suisse 78: 1-17.
- Nielsen, M.G., G.W. Elmes and V.E. Kipyatkov. 1999. Respiratory Q10 varies between populations of two species of Myrmica ants according to the latitude of their sites. Journal of Insect Physiology 45:559-564
- Noordijk, J., R. Morssinkhof, P. Boer, A. P. Schaffers, Th. Heijerman and K. V. Sýkora. 2008. How ants find each other; temporal and spatial patterns in nuptial flights. Insectes Sociaux 55(3):266-273.
- Nylander, W. "Synopsis des Formicides de France et d'Algérie." Annales des Sciences Naturelles, Zoologie (4)5 (1856): 51-109.
- O'Rourke F. J. 1948. The distribution and general ecology of the Irish Formicidae. Proceedings of the Royal Irish Academy. Section B: Biological, Geological, and Chemical Science 52: 383-410.
- Odegaard F. 2013. New and little known ants (Hymenoptera, Formicidae) in Norway. Norwegian Journal of Entomology 60, 172175.
- Ovazza M. 1953. Contributions a l'etude biologique de la Camargue , Formicides. Vie et Milieu 4(4) : 751-753.
- Palanca, A., A. Urbieta, C. Castán, and X. Espadaler. "Ants in the supraforestal pastures of the Tendeñera massif." Écologie des milieux montagnards et de haute altitude. Documents d'Ecologie Pyrénéenne III-IV (1984): 223-224.
- Paraschivescu D. 1978. Elemente balcanice in mirmecofauna R. S. Romania. Nymphaea 6: 463- 474.
- Park S.J., K.G. Kim, J.H. Kim, and B.J. Kim. 1998. Ants from the Seoraksan National Park. The Korean Journal of Systematic Zoology 14(4): 429-439.
- Passera L. 1967. Peuplement en fourmis terricoles du rebord Meridional des Causses jurassiques du Quercy: la lande calcaire a buis. Vie et milieu. Serie C, Biologie terrestre 18(1): 189-205.
- Paukkunen J., and M. V. Kozlov. 2015. Stinging wasps, ants and bees (Hy menoptera: Aculeata) of the Murmansk region, Northwest Russia. — Entomol. Fennica. 26: 53–73.
- Pavlova N. S. 2014. To ant fauna (Hymenoptera, Formicidae) of the floodplain of Medvedista River (Saratov Province). Entomological and parasitological studies in the Volga region 11: 145-147.
- Pech P., and K. Pruskova. 2013. Myrmica scabrinodis as a possible host of Myrmica hirsuta (Hymenoptera: Formicidae). Entomologica Fennica 24: 140-141.
- Petal J. 1974. Analysis of a sheep pasture ecosystem in the Pieniny mountains (the Carpathians) XV. The effect of pasture management on ant population. Ekologia Polska 22(3/4): 679-692.
- Petal J. 1994. Reaction of ant communities to degradations of forest habitats in the Karkonosze Mountains. Polish Academy of Sciences, Museum and Institute of Zoology, Memorabilia Zoologica 48: 171-179.
- Petal J. M. 1963. Faune des fourmis de la reserve de tourbiere en projet a Rakowskie Bagno pres de Frampol (voivodie de Lublin). Annales Universitatis Mariae Curie-Sk?odowska 58(7): 143-174.
- Petal J. M. 1967. Contribution a la connaissance des fourmis (Formicidae, Hymenoptera) de la region de Lublin. VII. Associations des fourmis des milieux de tourbieres, de forets et de dunes aux environs de Libiszow (dist. De Parczew). Annales Universitatis Mariae Curie-Sklodowska Lublin-Polonia 22(9): 117-130.
- Petal J. M. 1976. The effect of mineral fertilization on ant populations in meadows. Polish Ecological Studies 2(4): 209-218.
- Petal J., and A. Kusinska. 1994. Fractional composition of organic matter in the soil of anthills and of the environement of meadows. Pedobiologia 38: 493-501.
- Petrov I. Z. 2002. Contribution to the myrmecofauna (Formicidae, Hymenoptera) of the Banat Province (Serbia). Archives of Biological Sciences, Belgrade, 54(12): 57-64.
- Petrov I. Z., and C. A. Collingwood. 1992. Survey of the myrmecofauna (Formicidae, Hymenoptera) of Yugoslavia. Archives of Biological Sciences (Belgrade) 44: 79-91.
- Piers H. 1922. List of a small collection of ants (Formicidae) obtained in Queen's County, Nova Scotia, by the late Walter H. Prest. Proceedings and Transactions of the Nova Scotian Institute of Science, 15(4), 169-173.
- Poldi B., M. Mei, and F. Rigato. 1995. Hymenoptera, Formicidae. Vol 102. Checklist delle specie della fauna Italiana: 1-10.
- Punttila P., Y. Haila, J. Niemela, and T. Pajunen. 1994. Ant communities in fragments of old-growth taiga and managed surroundings. Ann. Zool. Fennici 31: 131-144.
- Pusvaskyte O. 1979. Myrmecofauna of the Lituanian SSR. Acta Entomologica Lituanica 4: 99-105.
- Radchenko A. G. 2007. The ants (Hymenoptera, Formicidae) in the collection of William Nylander. Fragmenta Faunistica (Warsaw) 50: 27-41.
- Radchenko A. G., and G. W. Elmes. 2003. A taxonomic revision of the socially parasitic Myrmica ants (Hymenoptera: Formicidae) of the Palaearctic region. Annales Zoologici (Warsaw) 53: 217-243.
- Radchenko A. G., and G. W. Elmes. 2010. Myrmica ants (Hymenoptera: Formicidae) of the Old World. Fauna Mundi 3. Warsaw: Natura Optima Dux Foundation, 790 pp.
- Reznikova Z. I. 2003. Distribution patterns of ants in different natural zones and landscapes in Kazakhstan and West Siberia along a meridian trend. Euroasian Entomological Journal 2(4): 235-342.
- Saaristo M. I. 1990. New provincial records for twelve ant species (Hymenoptera, Formicidae) from Finland. Entomologica Fennica 1: 191-192.
- Sadil J. V. 1952. A revision of the Czechoslovak forms of the genus Myrmica Latr. (Hym.). Sb. Entomol. Oddel. Nár. Mus. Praze 27: 233-278.
- Salata S. 2014. Ants (Hymenoptera: Formicidae) of the National Park of the Stołowe Mts. Przyroda Sudetow 17: 161-172.
- Salata S., and L. Borowiec. 2018. Taxonomic and faunistic notes on Greek ants (Hymenoptera: Formicidae). Annals of the Upper Silesian Museum in Bytom Entomology 27: 1-51.
- Salata S., and L. Borowiec. 2019. Preliminary division of not socially parasitic Greek Temnothorax Mayr, 1861 (Hymenoptera, Formicidae) with a description of three new species. ZooKeys 877: 81-131.
- Sanchez-Gil Jimeno R., and J. L. Reyes-Lopez. 2016. Study of ants species of the Sierra de San Carlos del Valle (Ciudad Real) and updating the provincial check list (Hymenoptera, Formicidae). Boln. Asoc. esp. Ent. 40 (1-2): 93-109.
- Santschi F. 1931. Notes sur le genre Myrmica (Latreille). Revue Suisse de Zoologie 38: 335-355.
- Santschi, F. "Notes sur les fourmis paléarctiques. II. Fourmis d'Asie Mineure récoltées par M. H. Gadeau de Kerville." Boletín de la Real Sociedad española de Historia natural (Madrid) 21 (1921): 110-116.
- Schlick-Steiner B. C., and F. M. Steiner. 1999. Faunistisch-ökologische Untersuchungen an den freilebenden Ameisen (Hymenoptera: Formicidae) Wiens. Myrmecologische Nachrichten 3: 9-53.
- Schlick-Steiner, B.C., F.M. Steiner, K. Moder, A. Bruckner, K. Fiedler and E. Christian. 2006. Assessing ant assemblages: pitfall trapping versus nest counting (Hymenoptera, Formicidae). Insectes Sociaux 53:274-281
- Seifert B. 1988. A taxonomic revision of the Myrmica species of Europe, Asia Minor, and Caucasia (Hymenoptera, Formicidae). Abhandlungen und Berichte des Naturkundemuseums Görlitz 62(3): 1-75.
- Seifert B. 1998. Rote Liste der Ameisen. - in: M. Binot, R. Bless, P. Boye, H. Gruttke und P. Pretscher: Rote Liste gefährdeter Tiere Deutschlands. Bonn-Bad Godesberg 1998: 130-133.
- Seifert B., A. Bagherian Yazdi, and R. Schultz. 2014. Myrmica martini sp n - a cryptic species of the Myrmica scabrinodis species complex (Hymenoptera: Formicidae) revealed by geometric morphometrics and nest-centroid clustering. Myrmecological News 19: 171-183.
- Seifert B., A. Bagherian Yazdi, and R. Schultz. 2014. Myrmica martini sp. n. - a cryptic species of the Myrmica scabrinodis species complex (Hymenoptera: Formicidae) revealed by geometric morphometrics and nest-centroid clustering. Myrmecological News 19: 171-183.
- Seifert, B. "Firm evidence for synonymy of Myrmica rugulosoides Forel, 1915 and Myrmica scabrinodis Nylander, 1846." Abhandlungen und Berichte des Naturkundemuseums Görlitz 58(6) (1984): 1-8.
- Seima F. A. 1964. On the fauna and ecology of ants in the Oka Preserve. Zoologicheskii Zhurnal 43: 1404-1408.
- Sellier Y., C. Galkowski, C. Lebas, and P. Wegnez. 2016. Découverte de Temnothorax pardoi (Tinaut, 1987) dans la réserve naturelle nationale du Pinail (Hymenoptera, Formicidae). Revue de l’Association Roussillonnaise d’Entomologie 25(2): 106-113.
- Shlyakhtenok A. S. 2007. Hymenoptera Aculeata of Raised Bogs in Belarus. Entomological Review 87(2): 136147.
- Skorka, P., M. Witek and M. Woyciechowski. 2006. A simple and nondestructive method for estimation of worker population size in Myrmica ant nests. Insectes Sociaux
- Slipinski P., M. Zmihorski, and W. Czechowski. 2012. Species diversity and nestedness of ant assemblages in an urban environment. Eur. J. Entomol. 109: 197206.
- Smith M. R. 1934. A list of the ants of South Carolina. Journal of the New York Entomological Society 42: 353-361.
- Sommer F. 1984. Etude de la myrmécofaune de la réserve de la Massane. Laboratoire Arago, Banyuls sur Mer (66), 29 pages.
- Sommer F., and H. Cagniant. 1988. Peuplements de fourmis des Albères Orientales (Pyrénées-Orientales, France) (Première partie). Vie Milieu 38: 189-200.
- Sonnenburg H. 2005. Die Ameisenfauna (Hymenoptera: Formicidae) Niedersachsens und Bremens. Braunschweiger Naturkundliche Schriften 7: 377-441.
- Steffek J., and M. Wiezik. 2008. Exploited peatbog and associated mollusk and ant assemblages: still a reasonable protection? Naturae Tutela 12: 15-19.
- Steiner F. M., S. Schödl, and B. C. Schlick-Steiner. 2002. Liste der Ameisen Österreichs (Hymenoptera: Formicidae), Stand Oktober 2002. Beiträge zur Entomofaunistik 3: 17-25.
- Stukalyuk S. V. 2015. Structure of the ant assemblages (Hymenoptera, Formicidae) in the broad-leaved forests of Kiev. Entomological Review 95(3): 370–387.
- Stukalyuk S. V. 2017. Stratification of the ant species (Hymenoptera, Formicidae) in the urban broadleaf woodlands of the city of Kiev. Entomological Review 97(3): 320-343.
- Stumper R. 1953. Etudes myrmecologiques. XI. Fourmis luxembourgeoises. Bulletin Soc. Nat. luxemb. 57: 122-135.
- Sushko G. G. 2013. The ants (hymenoptera, formicidae) bogs Belarusian Lakeland. Proceedings of the international. scientific-practical. conference, Vitebsk, 21-22 November 2013 - Vitebsk, 2013. - P. 215-216.
- Suvak M. 2007. The ants (Hymenoptera: Formicidae) in the stubble-fields of east Slovakia. Acta Zoologica Universitatis Comenianae. 47: 21-33
- Suñer i Escriche, David. "Contribució al coneixement mirmecologic de Gavarres, Montgrí, Guilleríes i la Serralada Transversal." Tesis Doctoral Universida (1991): 577 pp.
- Sveum P. 1979. Notes on the distribution of some Norwegian ant species (Hymenoptera, Formicidae). Fauna Norv. ser. B. 26: 10-11.
- Szujecki A., J. Szyszko, S. Mazur, and S. Perlinski. 1978. A succession of the ants (Formicidae) on afforested arable land and forest soils. Memorabilia Zoologica 29: 183-189.
- Tartally, A., D.R. Nash, S. Lengyel and Z. Varga. 2008. Patterns of host ant use by sympatric populations of Maculinea alcon and M. rebeli in the Carpathian Basin. Insectes Sociaux 55:370-381
- Tausan I., J. Dauber, M. R. Tricia, and B. Marko. 2017. Succession in ant communities (Hymenoptera: Formicidae) in deciduous forest clear-cuts – an Eastern European case study. European Journal of Entomology 114: 92-100.
- Tausan I., M. M. Jerpel, I. R. Puscasu, C. Sadeanu, R. E. Brutatu, L. A. Radutiu, and V. Giurescu. 2012. Ant fauna (Hymenoptera: Formicidae) of Sibiu County (Transylvania, Romania). Brukenthal. Acta Musei 7(3): 499-520.
- Tausan I., and B. Marko. 2009. Comparative analysis of ant communities (Hymenoptera: Formicidae) in the surroundings of Sibiu (Romania). Brukenthal. Acta Musei 4(3): 635-644.
- Tinaut A. 2016. Ants of the Tejeda, Almijara and Alhama Mountains Natural Park (Andalusia, Spain) (Hymenoptera, Formicidae). Boln. Asoc. esp. Ent., 40 (1-2): 125-159.
- Tinaut, A., J. Jiménez Rojas, and R. Pascual. "Estudio de la mirmecofauna de los bosques de Quercus Linneo 1753 de la provincia de Granada." Ecología 8 (1995): 429-438.
- Tizado Morales, E. J. Estudio comparado de la fauna y la biología de pulgones (Hom.), afidíinos (Hym.) y otros insectos acompañantes en dos áreas de la provincia de León In Secretariado de Publicaciones, Tesis doctoral en microficha, nº 67. León: Universidad de León, 1991.
- Urbieta Balado A., A. Palanca Soler, C. Castan Lanaspa, and X. Espadaler. 1984. Ants in the supraforestal pastures of the Tendenera massif. Documents dEcologie Pyrénéenne III-IV: 223-224.
- Vagalinski B., and A. Lapeva-Gjonova. 2012. The ants (Hymenoptera, Formicidae) of Plana Mountain (Bulgaria). Historia naturalis bulgarica 20: 87-101.
- Vankerkhoven F., Z. Vanstraelen, and W. Dekoninck. 2011. Rediscovery of the parasitic ant Myrmica karavajevi (Arnoldi, 1930) in Belgium (Formicidae, Hymenoptera). Bulletin S. R. B. E. 147: 185-187.
- Vepsalainen K., H. Ikonene, and M. J. Koivula. 2008. The structure of ant assembalges in an urban area of Helsinki, southern Finland. Ann. Zool. Fennici 45: 109-127.
- Vepsalainen K., R. Savolainen, J. Tiainen, and J. Vilen. 2000. Successional changes of ant assemblages: from virgin and ditched bogs to forests. Ann. Zool. Fennici 37: 135-149.
- Viereck H. L. 1903. Hymenoptera of Beulah, New Mexico. [part]. Trans. Am. Entomol. Soc. 29: 56-87.
- Vogrin, V. "Prilog fauni Hymenoptera - Aculeata Jugoslavije." Zast. Bilja 31(suppl.) (1955): 1-74.
- Weber N. A. 1948. A revision of the North American ants of the genus Myrmica Latreille with a synopsis of the Palearctic species. II. Annals of the Entomological Society of America 41: 267-308.
- Wegnez P. 2014. Premières captures de Lasius distinguendus Emery, 1916 et de Temnothorax albipennis (Curtis, 1854) au Grand-Duché de Luxembourg (Hymenoptera : Formicidae). Bulletin de la Société royale belge d’Entomologie 150 (2014) : 168-171.
- Wegnez P. 2017. Découverte de Myrmica lobicornis Nylander, 1846 et Lasius jensi Seifert, 1982, deux nouvelles espèces pour le Grand-Duché de Luxembourg (Hymenoptera: Formicidae). Bulletin de la Société royale belge d’Entomologie 153: 46-49.
- Wegnez P. 2018. Premières decouvertes de Myrmica bibikoffi Kutter, 1963 et de Ponera testacea Emery, 1895, au Luxembourg (Hymenoptera: Formicidae)Bulletin de la Société royale belge d’Entomologie 154: 263–272.
- Wegnez P., D. Ignace, E. Lommelen, M. Hardy, J. Bogaert, and C. Nilsson. 2015. Redécouverte de Teleutomyrmex schneideriKutter, 1950 dans les Alpes françaises (Hymenoptera: Formicidae). Bulletin de la Société royale belge d’Entomologie 151: 52-57.
- Wegnez P., and A. Ronk. 2017. Découverte de Camponotus herculeanus (Linnaeus, 1758) et signalement de quelques autres espèces rares de fourmis au Luxembourg (Hymenoptera : Formicidae). Bulletin de la Société des naturalistes luxembourgeois 119 : 153–159.
- Wegnez P., and F. Mourey. 2016. Formica uralensis Ruzsky, 1895 une espèce encore présente en France mais pour combien de temps ? (Hymenoptera: Formicidae). Bulletin de la Société royale belge d’Entomologie 152: 72-80.
- Wegnez P., and M. Fichaux. 2015. Liste actualisee des especes de fourmis repertoriees au Grand-Duche de Luxembourg (Hymenoptera: Formicidae). Bulletin de la Société royale belge d’Entomologie 151: 150-165
- Wheeler W. M. 1906. The ants of the Grand Cañon. Bulletin of the American Museum of Natural History 22: 329-345.
- Wheeler W. M. 1908. The ants of Casco Bay, Maine, with observations on two races of Formica sanguinea Latreille. Bulletin of the American Museum of Natural History 24: 619-645.
- Wheeler, William Morton. 1928. Ants of Nantucket Island, Mass. Psyche. 35(1):10-11.
- Wiezik M. 2008. Vertical distribution of ants from alpine habitats at Nízke Tatry mts. Naturae Tutela 12: 61-67.
- Wiezik M., A. Wiezikova, and J. Murin. 2008. Ant assemblages at horná orava peatbogs. Naturae Tutela 12: 55-60.
- Wiezik M., A. Wiezikova, and M. Svitok. 2010. Effects of secondary succession in abandoned grassland on the activity of ground-foraging ant assemblages (Hymenoptera: Formicidae). Acta Soc. Zool. Bohem. 74: 153-160
- Wiezik M., A. Wiezikova, and M. Svitok. 2011. Vegetation structure, ecological stability, and low-disturbance regime of abandoned dry grasslands support specific ant assemblages in Central Slovakia. Tuexenia 31: 301315.
- Wiezik M., M. Kozon, and A. Wiezikova. 2013. Ants of selected peat bog habitats at the Horná Orava protected area. Naturae Tutela 17(1): 57-63.
- Wieziková A. 2008. Epigaeic activity of ants (Hymenoptera: Formicidae) at hemerobic gradient: arable land green forage permanent grassland. Acta Facultatis Ecologiae 18: 69-76.
- Xia Yongjuan, and Zheng Zhemin. 1997. A survey of Formicidae form Xinjiang. Journal of Shaanxi Normal University 25(2): 64-66.
- Yazdi A. B., W. Münch, and B. Seifert. 2012. A first demonstration of interspecific hybridization in Myrmica ants by geometric morphometrics (Hymenoptera: Formicidae). Myrmecological News 17: 121-131.
- Yusopov Z. M. 2010. To the level of scrutiny of ants fauna (Hymenoptera, Formicidae from Kabardino-Balkarian state high-mountainous reserve (Central Caucasus). Proceedings of the Samara Scientific Center of RAS 12(5): 1464-1466.
- Zhuytszyuan D. 2016. The ants (Hymenoptera, Formicidae) Nizhne-Svirsky reserve and their environmental features. Master's thesis Saint Petersburg State University.
- Zryanin V. A., and T. A. Zryanina. 2007. New data on the ant fauna Hymenoptera, Formicidae in the middle Volga River Basin. Uspekhi Sovremennoi Biologii 127(2): 226-240.
- de Haro, Andrés, and C. A. Collingwood. "Prospección mirmecológica en la Cordillera Ibérica." Orsis 6 (1991): 129-126.
- Álvarez M., M. L. Munguira, and M. D. Martínez-Ibáñez. 2014. Comparative Study of the Morphology of Stridulatory Organs of the Iberian Lycaenid Butterfly Pupae (Lepidoptera). Journal of Morphology 275(4): 414430.
- Pages using DynamicPageList3 parser function
- Polygynous
- Invasive
- Photo Gallery
- North temperate
- FlightMonth
- Ant Associate
- Host of Myrmica karavajevi
- Host of Myrmica laurae
- Host of Myrmica vandeli
- Host of Lasius flavus
- ''Microdon'' fly Associate
- Host of Microdon myrmicae
- Phorid fly Associate
- Host of Pseudacteon brevicauda
- Aphid Associate
- Host of Aphis asclepiadis
- Host of Aphis forbesi
- Host of Brachycaudus helichrysi
- Host of Brachycaudus persicae
- Host of Cinara brevispinosa
- Host of Cryptomyzus ribis
- Host of Drepanaphis acerifoliae
- Ichneumonid wasp Associate
- Host of Hybrizon buccatus
- Butterfly Associate
- Host of Glaucopsyche alexis
- Host of Maculinea alcon
- Host of Maculinea teleius
- Fungus Associate
- Host of Rickia wasmanniii
- Nematode Associate
- Host of "Mermis"
- Host of Mermithidae (unspecified "Mermix")
- Cestode Associate
- Host of Davaineidae (Raillietina urogalli??)
- Host of Raillietina urogalli
- Karyotype
- Species
- Extant species
- Formicidae
- Myrmicinae
- Myrmicini
- Myrmica
- Myrmica scabrinodis
- Myrmicinae species
- Myrmicini species
- Myrmica species
- Ssr