Cardiocondyla minutior
| Cardiocondyla minutior | |
|---|---|

| |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Arthropoda |
| Class: | Insecta |
| Order: | Hymenoptera |
| Family: | Formicidae |
| Subfamily: | Myrmicinae |
| Tribe: | Crematogastrini |
| Genus: | Cardiocondyla |
| Species: | C. minutior |
| Binomial name | |
| Cardiocondyla minutior Forel, 1899
| |
| Synonyms | |
| |


{kind=link}
| Common Name | |
|---|---|
| Hime-hadaka-ari | |
| Language: | Japanese |
| Notes: | as Cardiocondyla tsukuyomi |
Believed to be native to the Indomalayan region (Seifert 2003), Cardiocondyla minutior is a pantropical tramp species. It is not known to be a pest in any area nor has it been known to cause harm to any native species in its introduced range. Dimorphic males, both alate and ergatoid, are known in this species.
| At a Glance | • Highly invasive • Ergatoid male |
Identification
A member of the Cardiocondyla minutior group.
C. minutior workers can be separated from Cardiocondyla emeryi by its lower profile postpetiole, a lack of an anteroventral bulge on the postpetiole, a lack or near lack of a metanotal groove, longer tergite pubescence, and more developed microsetae on eyes.
Seifert (2003) includes more details about specific morphological measures that separate these species. There is also a key to the holarctic species of Cardiocondyla that includes this and other tropical tramp species.
Sharaf et al. (2017) - Worker. Eyes small, with distinct microsetae; posterior margin of head straight or very weakly concave; anterior clypeal margin with feeble median concavity; mesosomal outline in profile nearly straight or feebly convex; metanotal groove weakly impressed or entirely absent; propodeal spines short and acute; petiole in profile with concave anterior face and rounded node; petiolar node in dorsal view circular and as long as broad; postpetiole low, with a completely flat sternite and without anteroventral bulge. Head, mesosoma, petiole and postpetiole varying from dirty yellow to dark dirty brown, gaster brown to blackish brown.
For the Australian fauna, Heterick (2019) provides the following notes: Cardiocondyla minutior was identified using the taxonomic keys to species-group and then to species in Seifert (2003). (Note: Seifert’s species level key bases the differentiation between C. minutior and its sister species Cardiocondyla tjibodana on minute morphometric differences and colour-tone, which, in practical terms, is rather unsatisfactory. Cardiocondyla minutior has a greater dispersal capacity and is a pantropical tramp species whereas C. tjibodana is mainly found on Indo-Malay and Pacific islands. The worker examined came from within the urban quarter of Darwin and is a dull yellowish-brown, so the former name is preferred. Seifert regarded the separation of the two taxa as tentative and conceded future investigations might reveal the two ants are conspecific. In such a case, C. minutior would be the senior name.)
Seifert (2022) provides the following details:
Worker: Small size, CS 422 µm. Head elongated, CL/CW 1.263. Postocular distance very large, PoOc/CL 0.475. Scape short, SL/CS 0.754. Eye rather small, EYE/CS 0.232, with notable micropilae, the longest measuring 6–10 µm. Occipital margin straight or very weakly concave. Frontal carinae slightly converging immediately caudal of FRS level (FL/FR 1.062). Dorsal profile of mesosoma rather straight or weakly convex, metanotal groove only suggested or entirely absent (MGr/CS 0.39%). Spines rather short and acute (SP/CS 0.133), their axis in profile deviating by 40–45° from longitudinal axis of mesosoma. Petiole rather wide and high (PeW/CS 0.283, PeH/CS 0.332), in profile with concave anterior face and rounded node that is in dorsal view circular and as long as wide. Postpetiole rather wide and very low (PpW/CS 0.475, PpH/CS 0.274), its sternite completely flat, without any anteroventral bulge; in dorsal view with angulate-convex sides and straight anterior margin. Paramedian vertex with deeply impressed, flat-bottomed foveolae of 13–18 µm diameter in dense honey-comb arrangement (if not displaced by longitudinal carinulae). Foveolae with an inner corona (margin of a flat tubercle) of 7–8 µm diameter. Mesosoma on whole surface sculptured. Waist segments with shallower and finer reticulum than on mesosoma, nodes sometimes slightly shiny. First gaster tergite with fine microreticulum that may be obscured by polluted surfaces. Pubescence on whole body long and dense, PLG/CS 7.9%, sqPDG 3.23. Color of head, mesosoma, and waist varying considerably from dirty yellowish to dark dirty brown, gaster dark to blackish brown.
Cardiocondyla minutior is phenotypically extremely similar to Cardiocondyla goa and Cardiocondyla tjibodana. Yet, the three taxa can be clustered by the exploratory data analyses (EDAs) NC-Ward, NC-part.hclust and NC-NMDS.kmeans using the 15 characters CS, CL/CW, PoOc/CL, SL/CS, FRS/CS, EYE/CS, MGr/CS, SP/CS, SpBa/CS, PeW/CS, PeH/CS, PpW/CS, PpH/CS, sqPDG and PLG/CS. Excluding single-specimen samples, a total of 79 samples with 194 worker specimens was available for analysis in these taxa. The EDA clustering was checked by running a controlling linear discriminant analysis (LDA) with species hypotheses allocated when the three EDAs agreed in classification but samples run as wild-cards when the EDAs classified ambiguously. The final species hypothesis established by the LDA determined the error on the sample level as 3.7% for NC-Ward and 6.2% for both NC-part.hclust and NC-NMDS.kmeans (Figure 28). Running the LDA with the final species hypothesis assigned to all 194 workers specimens, the classification error on the individual level was 3.6%. Naming the clusters in Figure 28 from left to right as clusters 1, 2 and 3 and running the specimens of the type series as wild-cards in the LDA, the following posterior probabilities were found on the sample level. The holotype series of C. tjibodana was allocated to cluster 1 with p = 0.9992. Allocation to cluster 2 was given with p = 0.9989 in the holotype series of C. minutior, with p = 0.9799 in the holotype series of C. tsukuyomi and with p = 0.8254 in the holotype series of C. breviscapus, hence the latter two taxa should represent junior synonyms of C. minutior. Allocation to cluster 3 was established with p = 0.9984 in the holotype of C. goa and with p = 0.9997 in the paratype series of C. goa.
A mean error of the three EDAs on the sample level of 5.4% is larger than the threshold of 4% as it was recommended by Seifert (2020). This might be a consequence of the small mean sample size (only 2.5 workers per supposed nest sample) and of a rather weak character system not sufficient to allow a better resolution. The phenotypical clustering is supported by differences in geographic distribution between the three taxa. Anyway, the critical error rate of the EDAs indicates a taxonomic conflict and the necessity to check the phenotypical clusters by an adequate system of nuclear DNA markers.
Keys including this Species
- Key to US Cardiocondyla species
- Key to Holartic Cardiocondyla
- Key to Micronesian Ants
- Key to Cardiocondyla of Hispaniola
- Key to selected species of Cardiocondyla
- Key to Palaearctic Cardiocondyla
Distribution
Worldwide distributed in the tropical zone but so far not confirmed for Africa. According to determined vouchers, distributed between 19° S and 33° N. The verified findings are indicated in the previous paragraph. Hypothesizing the Indo-Malayan region as natural range, it is the species with the largest tramp species potential within the C. minutior group, having colonized the Caribbean and diverse Pacific islands. It is intriguing that some 72% of all samples are from the latter two areas whereas only 16% are from the Indo-Malayan region (see also Wetterer, 2014). There are no reports on occurrence in greenhouses of the temperate zone. (Seifert, 2022)
Latitudinal Distribution Pattern
Latitudinal Range: 30° to 19°.
| North Temperate |
North Subtropical |
Tropical | South Subtropical |
South Temperate |
- Source: Seifert, 2022
Distribution based on Regional Taxon Lists
Afrotropical Region: Socotra Archipelago, Yemen.
Australasian Region: Australia, New Zealand.
Indo-Australian Region: Guam, Hawaii, Indonesia, Marshall Islands, Micronesia (Federated States of), New Guinea, Northern Mariana Islands.
Nearctic Region: United States.
Neotropical Region: Barbados, Costa Rica, Dominican Republic, Ecuador, French Guiana, Galapagos Islands, Greater Antilles, Grenada, Guatemala, Honduras, Mexico.
Oriental Region: India (type locality), Nepal, Sri Lanka, Vietnam.
Palaearctic Region: Japan, Oman.
Distribution based on AntMaps
Distribution based on AntWeb specimens
Check data from AntWeb
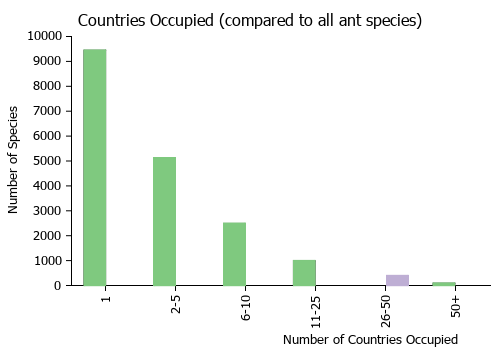
Countries Occupied
| Number of countries occupied by this species based on AntWiki Regional Taxon Lists. In general, fewer countries occupied indicates a narrower range, while more countries indicates a more widespread species. |
|
Estimated Abundance
| Relative abundance based on number of AntMaps records per species (this species within the purple bar). Fewer records (to the left) indicates a less abundant/encountered species while more records (to the right) indicates more abundant/encountered species. |

|
Biology
Seifert (2003): K. Yamauchi (pers. comm 2001) reported for Okinawa the nesting in shallow soil in open, disturbed areas with bare or weakly herbaceous ground. The Japanese population of C. minutior is reported to have a karyotype of 2n = 30 and to produce alate and ergatoid males. The latter perform lethal fighting for exclusive mating (Terayama 1999).
In Brazil, Tobago, and other areas, whereto the species has accidentally been introduced, nests can also be found in the soil, e.g., in open patches in gardens and plantations (Heinze pers. comm., reported in Seifert, 2022).
Both winged and ergatoid males occur. Winged males disperse, typically after mating within their natal colony. Ergatoid males are not known to disperse from their natal colony. Instead, they fight other ergatoid males, typically resulting in the death of all but one resident male (Heinze et al., 2004). As any Cardiocondyla species with tramp species potential, C. minutior is not known to have significant ecological impacts and it seems unlikely that it will ever become a significant pest after further range expansion. Ward (2005) reported that C. minutior was able to survive in sites dominated by the Argentine Ant Linepithema humile. (Seifert, 2022)
Male-male competition has been studied in this species. An abstract from one of these studies (Heinze et al. 2004): Wingless (ergatoid) males of the tramp ant Cardiocondyla minutior attack and kill their young ergatoid rivals and thus attempt to monopolize mating with female sexuals reared in the colony. Because of the different strength of local mate competition in colonies with one or several reproductive queens, we expected the production of new ergatoid males to vary with queen number. Sex ratios were mostly female-biased, but in contrast to the sympatric species C. obscurior (Cremer and Heinze, 2002) neither the percentage of ergatoid males nor of female sexuals among the first 20 sexuals produced varied considerably with queen number. As in C. obscurior, experimental colony fragmentation led to the production of winged males, whereas in unfragmented control colonies only ergatoid males eclosed.
Sarnat (2008) provides video of foraging workers.
Sharaf et al. (2017) provide the following notes from Yemen - The nesting and foraging behaviours of Cardiocondyla minutior are diverse. Workers were found in leaf litter under a date palm tree where the soil was moist and rich in accumulations of domestic livestock waste. Several specimens were observed foraging in leaf litter on a mountainside near a drainage where the soil was moist and supported diverse plant cover but was dominated by Adiantum capillus-veneris L. (Pteridaceae). A number of workers were found foraging in dry leaf litter under a small Fagonia tree (Zygophyllaceae). Several workers were found nesting in the moist basal leaves of a date palm tree trunk where hundreds of Hypoponera punctatissima workers co-occurred.
A specimen (as Cardiocondyla breviscapa from Oman was collected from "date palm" but it is not clear if this was taken in a date palm orchard or a specimen was found on an actual date palm (Phoenix dactylifera).
Castes
Worker
Images from AntWeb
   
| |
| Worker. Specimen code casent0103436. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by LACM, Los Angeles, CA, USA. |
   
| |
| Worker. Specimen code casent0103740. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by ABS, Lake Placid, FL, USA. |
   
| |
| Worker. Specimen code casent0173261. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by CDRS, Galapagos, Ecuador. |
   
| |
| Holotype of Cardiocondyla breviscapa. Worker. Specimen code casent0901759. Photographer Ryan Perry, uploaded by California Academy of Sciences. | Owned by NHMUK, London, UK. |
Queen
Images from AntWeb
   
| |
| Queen (alate/dealate). Specimen code casent0173260. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by CDRS, Galapagos, Ecuador. |
   
| |
| Queen (alate/dealate). Specimen code casent0178349. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by CAS, San Francisco, CA, USA. |
    
| |
| Queen (alate/dealate). Specimen code casent0103435. Photographer April Nobile, uploaded by California Academy of Sciences. | Owned by LACM, Los Angeles, CA, USA. |
Nomenclature
The following information is derived from Barry Bolton's Online Catalogue of the Ants of the World.
- minutior. Cardiocondyla nuda var. minutior Forel, 1899a: 120 (w.) HAWAII.
- Type-material: syntype workers (number not stated).
- Type-localities: Hawaiian Is (“Sandwich Is”): Oahu I., Honolulu, 1893 (R.C.L. Perkins), Hawaiian Is: Molokai I., Mts, 3000 ft, 1893 (R.C.L. Perkins).
- Type-depositories: BMNH, MHNG.
- Wheeler, W.M. 1922f: 317 (q.); Heinze, 1999: 251 (polymorphic m.).
- Subspecies of nuda: Forel, 1903d: 404; Forel, 1907a: 17; Wheeler, W.M. 1922f: 317; Emery, 1922e: 126; Wheeler, W.M. 1932a: 7; Wheeler, W.M. 1934h: 14; Wheeler, W.M. 1935g: 21; Smith, M.R. 1944a: 38 (redescription). Creighton, 1950a: 198; Smith, M.R. 1951a: 807; Smith, M.R. 1958c: 125; Smith, M.R. 1967: 355.
- Junior synonym of nuda: Wilson & Taylor, 1967: 55; Smith, D.R. 1979: 1375; Bolton, 1995b: 132; Zhou, 2001b: 87; Lyu & Cho, 2003b: 267 (error).
- Status as species: Heinze, 1999: 251; Seifert, 2003a: 283 (redescription); Ward, 2005: 65; Don, 2007: 85; Clouse, 2007b: 230; Terayama, 2009: 179; Branstetter & Sáenz, 2012: 257; Sarnat & Economo, 2012: 73; Ramage, 2014: 169; Wetterer, 2014b: 567; Bharti, Guénard, et al. 2016: 34; Wetterer, et al. 2016: 10; Sharaf, Fisher, et al. 2017: 20; Deyrup, 2017: 56; Dekoninck, et al. 2019: 1153; Lubertazzi, 2019: 97; Dias, R.K.S. et al. 2020: 62; Seifert, 2022b: 26 (diagnosis); Sharaf, Wetterer, et al. 2022: 41; Sharaf, Salman & Aldawood, 2023: 201; Herrera, Tocora, et al. 2024: 176.
- Senior synonym of breviscapus: Seifert, 2022b: 26.
- Senior synonym of tsukuyomi: Seifert, 2003a: 283; Seifert, B. 2022b: 26.
- Distribution [tramp species]
- Austral: New Caledonia, New Zealand.
- Malagasy: Aldabra, Chagos Archipelago, Europa I., Madagascar, Mayotte, Seychelles.
- Malesian: Fiji Is, French Polynesia, Hawaii, Indonesia (Biak), Kiribati (Line Is), Micronesia, Niue, Palau, Papua New Guinea (Bismarck Archipelago).
- Nearctic: U.S.A.
- Neotropical: Barbados, Brazil, Chile (Easter I.), Colombia, Dominican Republic, Ecuador (Galapagos Is), Guatemala, Puerto Rico, Trinidad (+ Tobago).
- Oriental: India (+ Nicobar Is), Japan, Nepal, Sri Lanka, Taiwan, Vietnam.
- Palaearctic: Egypt, Oman, Saudi Arabia, Yemen.
- breviscapus. Cardiocondyla breviscapus Seifert, 2003a: 288, fig. 64 (w.) INDIA (Tamil Nadu).
- Type-material: holotype worker, 2 paratype workers.
- Type-locality: holotype India: Coimbatore, 25.ix.1979 (J. Noyes); paratypes with same data.
- Type-depository: BMNH.
- Status as species: Bharti, Guénard, et al. 2016: 33; Sharaf, Fisher, et al. 2018: 18.
- Junior synonym of minutior: Seifert, 2022b: 26.
- tsukuyomi. Cardiocondyla tsukuyomi Terayama, 1999d: 101, figs. 11-13 (w.q.m. ergatoid m.) JAPAN.
- Type-material: holotype worker, 26 paratype workers, 7 paratype queens, 3 paratype males.
- Type-locality: holotype Japan: Okinawa Pref., Okinawa-jima, Ada, vi.1988 (K. Yamauchi); paratypes: 19 workers, 4 queens, 10 males with same data, 7 workers, 3 queens Okinawa Pref., Miyako Is, Kuruma-jima, 2.i.1984 (H. Takamine).
- Type-depositories: MNHA (holotype); MNHA, SMNG (paratypes).
- Status as species: Imai, et al. 2003: 153.
- Junior synonym of minutior: Seifert, 2003a: 283; Seifert, 2022b: 26.
Unless otherwise noted the text for the remainder of this section is reported from the publication that includes the original description.
Seifert (2003): Intraspecific variability in C. minutior is rather low within the huge range of its distribution, extending over the Neotropic, Polynesian, Australasian, Indo-Australasian, and Oriental regions. Samples from central Sri Lanka have significantly smaller eyes, those from Okinawa, N India, and Nepal shorter heads but all these deviating populations are in the vast majority of other characters consistent with the overall average.
Cardiocondyla tsukuyomi is in body shape and any structural and morphometric character consistent with the worldwide population of C. minutior as it is with the types of C. minutior from Hawaii. The 3 studied type workers of C. tsukuyomi and 4 topotypical non-type workers from Okinawa do not differ from the C. minutior population from outside Okinawa (Tab. 5). Recent mDNA studies have shown that tsukuyomi and minutior cluster closely together, while Cardiocondyla tjibodana and C. minutior could represent separate evolutionary lines (Trindl & Heinze, pers. comm. October 2002).
Description
Worker
Species redescribed in diagnosis, Seifert 2022 (see the identification section, above, of this species page). Cardiocondyla species treated in this revision are differentiated using exploratory data analyses and principle component analysis (Numeric Morphology-Based Alpha-Taxonomy/NUMOBAT) of worker measurements (Table S1 and S2, Seifert 2022 supplementary information).
Seifert (2003) - Small size, CS 418. Head elongated, CL/CW 1.259. Postocular distance very large, PoOC/CL 0.475. Scape short, SL/CS 0.756. Eye rather small, EYE 0.233, with notable microsetae, the longest measuring 6 - 10 µm. Occipital margin straight or very weakly concave. Frontal carinae slightly converging immediately caudal of FRS level. Anterior clypeal margin with suggested median concavity. Clypeus, frontal laminae, frontal triangle, and very narrow anteromedian stripe of vertex longitudinally carinulate (in some specimens from N India and Nepal such carinulae cover the whole median and paramedian vertex, with reduction of foveolae in these areas). Except for longitudinal rugae on preocular surface, semicircular rugae around antennal fossae, and few short, longitudinal rugae on metapleuron; whole body without any rugosity. Sculpture on paramedian vertex similar to situation in the C. emeryi types, showing deeply impressed, flat-bottomed foveolae of 13 - 18 µm diameter in dense honey-comb arrangement (if not displaced by longitudinal carinulae). Foveolae with an inner corona (margin of a f1at tubercle) of 7 - 8 µm diameter. Mesosoma on whole surface sculptured, rather mat: dorsal mesosoma densely and strongly reticulate-foveolate; lateral mesosoma densely and strongly reticulate; metapleuron with 1 - 4 longitudinal rugae. Waist segments with fully developed, but shallower and finer reticulum than on mesosoma, nodes sometimes slightly shining. First gaster tergite often with very fine microreticulum. Pubescence on whole body long and dense, sqrtPDG 3.34. Dorsal profile of mesosoma rather straight or weakly convex, metanotal groove only suggested or entirely absent. Spines short and acute, their axis in profile deviating by 40 - 45° from longitudinal axis of mesosoma. Petiole in profile with concave anterior face and rounded node that is in dorsal view circular and as long as wide. Postpetiole very low, its sternite completely fiat, without any anteroventral bulge; in dorsal view with angulate-convex sides and straight anterior margin. Colour of head, mesosoma, and waist varying considerably from dirty yellowish to dark dirty brown, gaster dark to blackish brown. For morphometric data of 72 workers see Tab. 14.
Queen
Seifert (2003) - Very small size. Head elongated, CL/CW 1.228. Scape rather short, SL/CS 0.755. Postocular index very large, PoOc/CL 0.459. Eyes with numerous hairs, the longest of them 8 - 11 µm long. Occipital margin more or less straight. Anteromedian clypeal margin straight to slightly convex. Vertex with deeply impressed, flat-bottomed, densely-packed foveolae of 15 - 18 µm diameter which show an inner corona of 7 -9 µm diameter. Paramedian and median areas of vertex with suggested longitudinal rugae. Frontal laminae and clypeus with few longitudinal carinulae. Whole dorsal area of mesosoma with deep, densely-packed foveolae, fragments of longitudinal rugae visible on mesonotum. Lateral lobes of praescutellum connected by a very thin junction or entirely separated. Lateral area of mesosoma foveolate-reticulate, region of metapleural gland bulla with longitudinal rugae. Propodeal spines well-developed, their axis deviating from mesosomal axis in lateral view by 25 -30°. Petiole node foveolate-reticulate, in dorsal view circular. Postpetiole in dorsal view strongly foveolate-reticulate, distinctly wider than long, with straight anterior margin and strongly convex sides. Postpetiolar sternite very flat, without any bulge. Whole body covered by long and dense pubescence. Dorsum of gaster shining, with fine microreticulum. For morphometric data of 14 gynes see Tab. 19.
Type Material
Seifert (2003):
Cardiocondyla minutior: two syntype workers labelled "C. nuda Mayr v. minutior type Forel, Hawai" and "Molockai Mts., 3000 ft. Perkins 1893", MHN Genève.
Cardiocondyla tsukuyomi: 6 paratype workers from the same sample as holotype, labelled: "VI 1988 K.Yamauchi leg., Ada, Okinawa-jima Okinawa Pref." and "Cardiocondyla tsukuyomi Terayama, 1999, Paratype", SMN Garlitz.
Cardiocondyla nuda var. minutior Forel, 1899; Hawaii: Honolulu and Molockai [types investigated].
Cardiocondyla tsukuyomi Terayama, 1999; Ada / Okinawa Island [types investigated], syn.n
Cardiocondyla breviscapa 3 syntype workers labelled “INDIA: Coimbatorc 25.ix.79 J. Noyes”, The Natural History Museum London. [Coimbatore is situated at 11.00 N, 76.58 E].
Karyotype
- See additional details at the Ant Chromosome Database.
 Explore: Show all Karyotype data or Search these data. See also a list of all data tables or learn how data is managed.
Explore: Show all Karyotype data or Search these data. See also a list of all data tables or learn how data is managed.
- 2n = 27 (Japan) (Imai & Yamauchi, unpublished (Japanese Ant Image Database)).
- 2n = 30 (Japan) (Terayama, 1999).
References
- Albuquerque, E., Prado, L., Andrade-Silva, J., Siqueira, E., Sampaio, K., Alves, D., Brandão, C., Andrade, P., Feitosa, R., Koch, E., Delabie, J., Fernandes, I., Baccaro, F., Souza, J., Almeida, R., Silva, R. 2021. Ants of the State of Pará, Brazil: a historical and comprehensive dataset of a key biodiversity hotspot in the Amazon Basin. Zootaxa 5001, 1–83 (doi:10.11646/zootaxa.5001.1.1).
- Baena, M.L., Escobar, F., Valenzuela, J.E. 2019. Diversity snapshot of green–gray space ants in two Mexican cities. International Journal of Tropical Insect Science 40, 239–250 (doi:10.1007/s42690-019-00073-y).
- Baer, B. 2011. The copulation biology of ants (Hymenoptera: Formicidae). Myrmecological News 14: 55-68.
- Brassard, F., Leong, C.-M., Chan, H.-H., Guénard, B. 2021. High diversity in urban areas: How comprehensive sampling reveals high ant species richness within one of the most urbanized regions of the world. Diversity 13, 358 (doi:10.3390/d13080358).
- Cantone S. 2018. Winged Ants, The queen. Dichotomous key to genera of winged female ants in the World. The Wings of Ants: morphological and systematic relationships (self-published).
- Creighton, W. S. 1950a. The ants of North America. Bulletin of the Museum of Comparative Zoology 104: 1-585 (page 198, Subspecies of nuda)
- Dekoninck, W., Wauters, N., Delsinne, T. 2019. Capitulo 35. Hormigas invasoras en Colombia. Hormigas de Colombia.
- Forel, A. 1899a. Heterogyna (Formicidae). Fauna Hawaii. 1: 116-122 (page 120, worker described)
- Franco, W., Ladino, N., Delabie, J.H.C., Dejean, A., Orivel, J., Fichaux, M., Groc, S., Leponce, M., Feitosa, R.M. 2019. First checklist of the ants (Hymenoptera: Formicidae) of French Guiana. Zootaxa 4674, 509–543 (doi:10.11646/zootaxa.4674.5.2).
- Gratiashvili, N., Kuschel, L., Heinze, J. 2020. Morphometry and colony structure of ants of the genus Cardiocondyla (Hymenoptera: Formicidae) from Georgia. Zoology in the Middle East 66, 347–356 (doi:10.1080/09397140.2020.1835216).
- Heinze J. 1999. Male polymorphism in the ant species Cardiocondyla minutior (Hymenoptera: Formicidae). Entomol. General. 23: 251-258. (page 251, polymoprhic male described)
- Heinze, J., Böttcher, A., Cremer, S. 2004. Production of winged and wingless males in the ant, Cardiocondyla minutior. Insectes Sociaux 51, 275–278.
- Heinze, J., S. Cremer, N. Eckl, and A. Schrempf. 2006. Stealthy invaders: the biology of Cardiocondyla tramp ants. Insectes Sociaux. 53:1-7. DOI: 10.1007/s00040-005-0847-4
- Heinze, J., Trindl, A., Seifert, B., Khin Ma Ma, Maung, W. 2020. First records of Cardiocondyla ants from Myanmar. Asian Myrmecology 12, e012005 (doi:10.20362/am.012005).
- Heinze, J.; Böttcher, A.; Cremer, S. 2004. Production of winged and wingless males in the ant, Cardiocondyla minutior. Insectes Soc. 51: 275-278. (page 275, see also)
- Herrera, H.W., Baert, L., Dekoninck, W., Causton, C.E., Sevilla, C.R., Pozo, P., Hendrickx, F. 2020. Distribution and habitat preferences of Galápagos ants (Hymenoptera: Formicidae). Belgian Journal of Entomology, 93: 1–60.
- Heterick, B.E. 2019. First record of the formicine genus Overbeckia (Hymenoptera: Formicidae) from Australia. Myrmecological News 29: 163-166 (doi:10.25849/myrmecol.news_029:163).
- Jacobs, S. 2020. Population genetic and behavioral aspects of male mating monopolies in Cardiocondyla venustula (Ph.D. thesis).
- Lubertazzi, D. 2019. The ants of Hispaniola. Bulletin of the Museum of Comparative Zoology, 162(2), 59-210 (doi:10.3099/mcz-43.1).
- MacGown, J.A., Booher, D., Richter, H., Wetterer, J.K., Hill, J.G. 2021. An updated list of ants of Alabama (Hymenoptera: Formicidae) with new state records. Transactions of the American Entomological Society 147: 961-981 (doi:10.3157/061.147.0409).
- Meurgey, F. 2020. Challenging the Wallacean shortfall: A total assessment of insect diversity on Guadeloupe (French West Indies), a checklist and bibliography. Insecta Mundi 786: 1–183.
- Rosas-Mejía, M., Guénard, B., Aguilar-Méndez, M. J., Ghilardi, A., Vásquez-Bolaños, M., Economo, E. P., Janda, M. 2021. Alien ants (Hymenoptera: Formicidae) in Mexico: the first database of records. Biological Invasions 23(6), 1669–1680 (doi:10.1007/s10530-020-02423-1).
- Sarnat, E.M. (2008) PIAkey: Identification guide to ants of the Pacific Islands, Edition 2.0, Lucid v. 3.4. USDA/APHIS/PPQ Center for Plant Health Science and Technology and University of California — Davis. [1]
- Satria, R. 2017. Taxonomy of the ant genus Odontomachus (Hymenoptera: Formicidae: Ponerinae) in the Indo-Chinese and Indo-Malayan subregions. Ph.D. thesis, Tokyo Metropolitan University.
- Seifert, B. 2003a. The ant genus Cardiocondyla (Insecta: Hymenoptera: Formicidae) - a taxonomic revision of the C. elegans, C. bulgarica, C. batesii, C. nuda, C. shuckardi, C. stambuloffii, C. wroughtonii, C. emeryi, and C. minutior species groups. Ann. (page 283, Senior synonym of tsukuyomi, raised to species)
- Seifert, B. 2022. The ant genus Cardiocondyla (Hymenoptera: Formicidae): The species groups with Oriental and Australasian origin. Diversity 15, 25 (doi:10.3390/d15010025).
- Seifert, B. 2023. A revision of the Palaearctic species of the ant genus Cardiocondyla Emery 1869 (Hymenoptera: Formicidae). Zootaxa 5274(1), 1–64 (doi:10.11646/zootaxa.5274.1.1).
- Sharaf, M. R., Wetterer, J. K., Mohamed, A. A., Aldawood, A. S. 2022. Faunal composition, diversity, and distribution of ants (Hymenoptera: Formicidae) of Dhofar Governorate, Oman, with updated list of the Omani species and remarks on zoogeography. European Journal of Taxonomy 838: 1-106 (doi:10.5852/ejt.2022.838.1925).
- Sharaf, M.R., Al Dhafer, H.M., Abdel-Dayem, M.S., Aldawood, A.S. 2024. Cardiocondyla hashemi sp. n., a new species of the C. batesii species-group (Hymenoptera: Formicidae) from Saudi Arabia, with a key to the Saudi species. Zoology in the Middle East]] (doi:10.1080/09397140.2024.2321640).
- Sharaf, M.R., Fisher, B.L., Collingwood, C.A., Aldawood, A.S. 2017. Ant fauna (Hymenoptera: Formicidae) of the Socotra Archipelago (Yemen): zoogeography, distribution and description of a new species. Journal of Natural History 51, 317–378 (DOI 10.1080/00222933.2016.1271157).
- Smith, J. H.; Atherton, D. O. 1944. Seed-harvesting and other ants in the tobacco-growing districts of North Queensland. Qld. J. Agric. Sci. 1(3 3: 33-61 (page 38, Subspecies of nuda)
- Subedi, I.P., Budha, P.B., Bharti, H., Alonso, L. 2020. An updated checklist of Nepalese ants (Hymenoptera, Formicidae). ZooKeys 1006, 99–136 (doi:10.3897/zookeys.1006.58808).
- Varela-Hernández, F., Medel-Zosayas, B., Martínez-Luque, E.O., Jones, R.W., De la Mora, A. 2020. Biodiversity in central Mexico: Assessment of ants in a convergent region. Southwestern Entomologist 454: 673-686.
- Ward, P.S. 2005. A synoptic review of the ants of California (Hymenoptera: Formicidae). Zootaxa 936, 1–68.
- Wetterer, J.K. 2014. Worldwide spread of the Lesser Sneaking ant, Cardiocondyla minutior (Hymenoptera: Formicidae). Florida Entomologist 567-574.
- Wetterer, J.K. 2017. Invasive ants of Bermuda revisited. Journal of Hymenoptera Research 54, 33–41 (doi:10.3897/jhr.54.11444).
- Wetterer, J.K. 2021. Ants (Hymenoptera, Formicidae) of St. Vincent, West Indies. Sociobiology 68, e6725 (doi:10.13102/sociobiology.v68i2.6725).
- Wheeler, W. M. 1922n. Formicidae from Easter Island and Juan Fernandez. Pp. 317-319 in: Skottsberg, C. (ed.) 1921-1940. The natural history of Juan Fernandez and Easter Island. Vol. III. Zoology. Uppsala: Almqvist & Wiksells, 688 pp. (page 317, queen described)
- Wilson, E. O.; Taylor, R. W. 1967b. The ants of Polynesia (Hymenoptera: Formicidae). Pac. Insects Monogr. 14: 1-109 (page 55, Junior synonym of nuda)
- Yamauchi, K., Y. Asano, B. Lautenschläger, A. Trindl, and J. Heinze. 2005. A new type of male dimorphism with ergatoid and short-winged males in Cardiocondyla cf. kagutsuchi. Insectes Sociaux. 52:274-281. DOI: 10.1007/s00040-005-0803-3
- Yoshizawa, J., K. Yamauchi, and K. Tsuchida. 2011. Decision-making conditions for intra- or inter-nest mating of winged males in the male-dimorphic ant Cardiocondyla minutior. Insectes Sociaux. 58:531-538. DOI: 10.1007/s00040-011-0175-9
- Yu, Y. 2016. Risk of alien species introduction to Ogasawara Islands : Case study of ants at Tokyo Port. World Heritage Studies 1, 86-89.
References based on Global Ant Biodiversity Informatics
- Eguchi K.; Bui T. V.; Yamane S. 2011. Generic synopsis of the Formicidae of Vietnam (Insecta: Hymenoptera), part I Myrmicinae and Pseudomyrmecinae. Zootaxa 2878: 1-61.
- Forel A. 1903. Les fourmis des îles Andamans et Nicobares. Rapports de cette faune avec ses voisines. Rev. Suisse Zool. 11: 399-411.
- Guénard B., and R. R. Dunn. 2012. A checklist of the ants of China. Zootaxa 3558: 1-77.
- Leong C. M., S. F. Shiao, and B. Guenard. 2017. Ants in the city, a preliminary checklist of Formicidae (Hymenoptera) in Macau, one of the most heavily urbanized regions of the world. Asian Myrmecology 9: e009014.
- Liu K. L., M. H. Peng, Y. C. Hung, and K. B. Neoh. 2019. Effects of park size, peri-urban forest spillover, and environmental filtering on diversity, structure, and morphology of ant assemblages in urban park. Urban Ecosystems https://doi.org/10.1007/s11252-019-00851-z)
- Seifert B. 2003. The ant genus Cardiocondyla (Insecta: Hymenoptera: Formicidae) - a taxonomic revision of the C. elegans, C. bulgarica, C. batesii, C. nuda, C. shuckardi, C. stambuloffii, C. wroughtonii, C. emeryi, and C. minutior species groups. Annalen des Naturhistorischen Museums in Wien. B, Botanik, Zoologie 104: 203-338.
- Terayama M. 2009. A synopsis of the family Formicidae of Taiwan (Insecta: Hymenoptera). Research Bulletin of Kanto Gakuen University. Liberal Arts 17:81-266.
- Terayama M., S. Kubota, and K. Eguchi. 2014. Encyclopedia of Japanese ants. Asakura Shoten: Tokyo, 278 pp.
- Terayama Mamoru. 2009. A synopsis of the family Formicidae of Taiwan (Insecta, Hymenoptera). The Research Bulletin of Kanto Gakuen University 17: 81-266.
- Terayama, M. 2009. A synopsis of the family Formicidae of Taiwan (Insecta; Hymenoptera). The Research Bulletin of Kanto Gakuen University 17: 81-266.
- Wetterer J. 2014. Worlwide spread of the lesser sneaking ant, Cardiocondyla minutior (Hymenoptera: Formicidae). Florida Entomologist 97(2): 567-574.