Veromessor chamberlini
| Veromessor chamberlini | |
|---|---|

| |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Arthropoda |
| Class: | Insecta |
| Order: | Hymenoptera |
| Family: | Formicidae |
| Subfamily: | Myrmicinae |
| Tribe: | Stenammini |
| Genus: | Veromessor |
| Species: | V. chamberlini |
| Binomial name | |
| Veromessor chamberlini (Wheeler, W.M., 1915)
| |


Little is known about the biology of V. chamberlini. Workers are diurnal and forage in a column (M. Bennett, pers. comm.). Colonies are small with a maximum of 627 workers (M. Bennett, pers. comm.). Workers are monomorphic. (Johnson et al., 2022)
Identification
Worker
This species is uniquely characterized by the following combination of features(Johnson et al., 2022):
- Yellowish-red to dark orangish-red
- Medial lobe of clypeus with two strong submedial rugae, sometimes with one to two weaker sublateral rugae, medial lobe not thick and protuberant in profile, not elevated above lateral lobes in frontal view
- Mandibles with 8 teeth
- Dorsal base of scape flattened and widened; maximum basal width of scape distinctly greater than maximal preapical width; in frontal view, base of scape asymmetrical with ventral portion of anterior edge greatly expanded compared to dorsal edge
- MOD less (usually much less) than OMD, OI < 22.0
- Cephalic dorsum with prominent wavy to weakly irregular rugae, medial rugae diverging toward posterior corners; rugae along posterior margin strongly irregular to weakly rugoreticulate; cephalic interrugae weakly to moderately coriarious, moderately shining
- Psammophore moderately well developed; ventral surface of head capsule with J-shaped hairs arranged in a V-shaped row which does not reach the posterior part of lateroventral margin of head capsule, usually mixed with straight or evenly curved hairs
- Dorsum of pronotum with weakly to strongly irregular rugae; sides of pronotum with widely spaced, regular to wavy, longitudinal rugae; mesonotum with irregular longitudinal rugae; mesopleura with regular to wavy rugae that traverse longitudinally to posterodorsally; interrugae on mesosoma weakly coriarious, shining
- Propodeal spines slender, acuminate, weakly curved posterad in profile and weakly curved inward in dorsal view; length > 3.0× the distance between their bases and length > 1.0× MOD; infraspinal facet and propodeal declivity weakly coriarious to weakly rugose, strongly shining
- Metasternal process moderately large, longer than high, apex broadly rounded, margin irregular and translucent (Figures 6C, 11, 12A)
 Johnson et al, 2022, Fig. 6. Photographs of the four categories of increasing psammophore development for species of Veromessor based on number and distribution of long J-shaped hairs on the ventral surface of the head capsule (= hypostomal region) (see text). Photograph of: (A) V. chicoensis (CASENT0923125)—J-shaped hairs mostly absent with scattered straight or evenly curved hairs, (B) V. andrei (CASENT0923140)—J-shaped hairs present but not arranged in a distinct row, usually mixed with straight or evenly curved hairs, (C) V. chamberlini (CASENT0761101) and (D) V. smithi (CASENT0923131)—J-shaped hairs present, arranged in a V-shaped row which does not reach the posterior part of the lateroventral margin of head capsule, usually mixed with straight or evenly curved hairs, and (E) V. pergandei (CASENT0923124)—J-shaped hairs present, many long J-shaped hairs arranged in a distinct row around the outer margin of the ventral region of the head capsule. Photographs by Wade Lee from www.AntWeb.org.
Johnson et al, 2022, Fig. 6. Photographs of the four categories of increasing psammophore development for species of Veromessor based on number and distribution of long J-shaped hairs on the ventral surface of the head capsule (= hypostomal region) (see text). Photograph of: (A) V. chicoensis (CASENT0923125)—J-shaped hairs mostly absent with scattered straight or evenly curved hairs, (B) V. andrei (CASENT0923140)—J-shaped hairs present but not arranged in a distinct row, usually mixed with straight or evenly curved hairs, (C) V. chamberlini (CASENT0761101) and (D) V. smithi (CASENT0923131)—J-shaped hairs present, arranged in a V-shaped row which does not reach the posterior part of the lateroventral margin of head capsule, usually mixed with straight or evenly curved hairs, and (E) V. pergandei (CASENT0923124)—J-shaped hairs present, many long J-shaped hairs arranged in a distinct row around the outer margin of the ventral region of the head capsule. Photographs by Wade Lee from www.AntWeb.org. Johnson et al., 2022, Fig. 12. Photographs of dorsal surface of scape: (A) Veromessor chamberlini (LACMENT359743)—anterior edge of scape curving inward distad of base (red arrow), posterior edge of scape flat to weakly convex at base (black arrow), narrowing distad of base; widened area at base of scape much longer than width at base, and (B) V. lobognathus (LACMENT363932)—anterior edge of scape flat at base (red arrow), posterior edge of scape narrowing from base (black arrow); widened area at base of scape shorter than width at base. Photographs by Michele Esposito from www.AntWeb.org.
Johnson et al., 2022, Fig. 12. Photographs of dorsal surface of scape: (A) Veromessor chamberlini (LACMENT359743)—anterior edge of scape curving inward distad of base (red arrow), posterior edge of scape flat to weakly convex at base (black arrow), narrowing distad of base; widened area at base of scape much longer than width at base, and (B) V. lobognathus (LACMENT363932)—anterior edge of scape flat at base (red arrow), posterior edge of scape narrowing from base (black arrow); widened area at base of scape shorter than width at base. Photographs by Michele Esposito from www.AntWeb.org.
,_Fig._6.jpg)
,_Fig._11.jpg)
,_Fig._12.jpg)
Queen
This caste is diagnosed by the following combination of features (Johnson et al., 2022):
- Orangish to orangishbrown
- Medial lobe of clypeus with two strong submedial rugae, sometimes with one to two weaker sublateral rugae, medial lobe not thick and protuberant in profile, not elevated above lateral lobes in frontal view
- Mandibles with 8 teeth
- Dorsal base of scape flattened and widened; maximum basal width of scape distinctly greater than maximal preapical width; in frontal view, base of scape asymmetrical with ventral portion of anterior edge greatly expanded compared to dorsal edge
- MOD similar to OMD
- Cephalic dorsum with prominent wavy to weakly irregular rugae, medial rugae diverging toward posterior corners; rugae along posterior margin weakly irregular to weakly rugoreticulate; interrugae on cephalic dorsum weakly to moderately coriarious, weakly shining,
- Psammophore moderately well developed
- Mesoscutum shining with fine longitudinal rugae, midline broadly smooth and shining between fine longitudinal rugae
- Propodeal spines shorter than distance between their bases; infraspinal facet smooth and shining
- Metasternal process moderately large, longer than high, apex broadly rounded, margin irregular and translucent (Figure 13)
,_Fig._13.jpg)
Male
This caste is diagnosed by the following combination of features (Johnson et al., 2022):
- Blackish brown
- Anepisternum densely lineopunctate; katepisternum strongly coriarious, anterior one-half of katepisternum with scattered, long, flexuous hairs, posterior one-half lacking long flexuous hairs except for a row of short hairs along posterior border
- Propodeum with widely spaced longitudinal rugae, interrugae densely punctate, dull; spines absent
- Metasternal process triangular, acute, with lamella extending on each side, portion posterior to lamella longer than portion anterior to lamella
- Subpetiolar process bluntly triangular, apex acute (Figures 1B, 14)
 Johnson et al., 2022, Fig. 1. Photographs of the metasternal process on males of Veromessor: (A) V. andrei (CASENT4010823), (B) V. chamberlini (UCRC_ENT00500152), (C) V. chicoensis (CASENT0869853), (D) V. julianus (LACMENT359792), (E) V. lariversi (CASENT0761204), (F) V. lobognathus (LACMENT363986), (G) V. pergandei (CASENT0869850), (H) V. pseudolariversi (CASENT0869851), (I) V. smithi (LACMENT364071), and (J) V. stoddardi (LACMENT364102). Photographs by Robert Johnson from www.AntWeb.org.
Johnson et al., 2022, Fig. 1. Photographs of the metasternal process on males of Veromessor: (A) V. andrei (CASENT4010823), (B) V. chamberlini (UCRC_ENT00500152), (C) V. chicoensis (CASENT0869853), (D) V. julianus (LACMENT359792), (E) V. lariversi (CASENT0761204), (F) V. lobognathus (LACMENT363986), (G) V. pergandei (CASENT0869850), (H) V. pseudolariversi (CASENT0869851), (I) V. smithi (LACMENT364071), and (J) V. stoddardi (LACMENT364102). Photographs by Robert Johnson from www.AntWeb.org.
,_Fig._1.jpg)
,_Fig._14.jpg)
Identification Notes
Veromessor chamberlini appears to have a relatively small geographic range (Johnson et al., 2022), such that it might be sympatric only with Veromessor andrei and Veromessor stoddardi. Workers of V. chamberlini are separated from both species based on: (1) moderately well developed psammophore, and (2) posterior margin of frontal lobes notably constricted. Both V. andrei and V. stoddardi have: (1) poorly developed psammophore, and (2) posterior margin of frontal lobes lacking a constriction.
A molecular phylogeny that used UCEs shows V. chamberlini and V. andrei are sister lineages (M.L Borowiec, unpub. data, in Johnson et al., 2022).
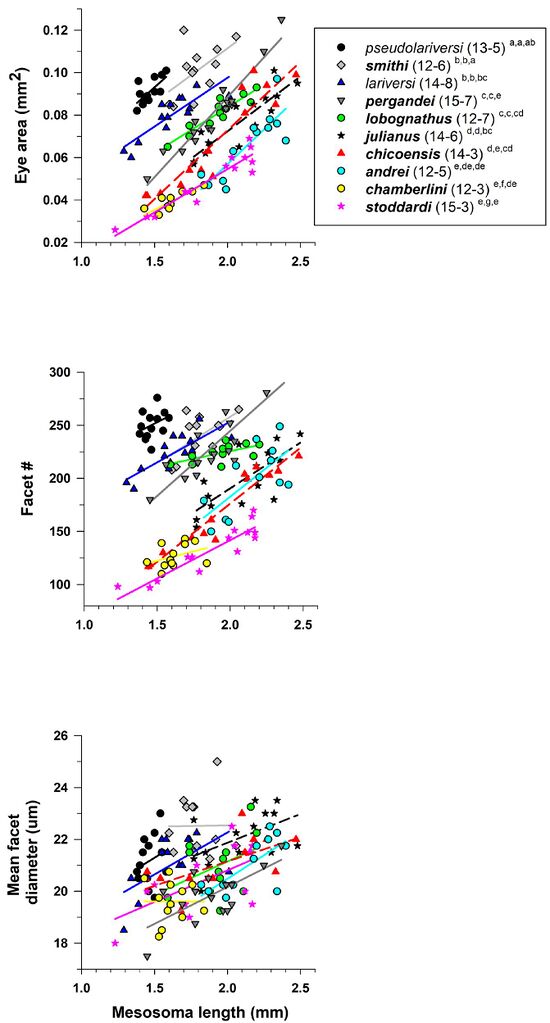 Johnson et al., 2022, Fig. 53. Eye area (mm2) (A), facet number (B), and mean facet diameter (μm) (C) for pale and dark colored species of Veromessor. Two species are pale (V. lariversi, V. pseudolariversi—open symbols and regular font), while the other eight species are dark (filled symbols and bold font). For each species, number of workers examined and number of colonies they were derived from is given in parentheses. Significant differences (P < 0.05) among species are denoted after each species name by the letters a–g: a > b > c > d > e > f > g; the three sets of letters for each species correspond to panels A, B, and C, respectively. Groupings are based on univariate F tests within MANCOVA followed by pairwise comparisons using a least significant differences test (see also Johnson & Rutowki, 2022).
Johnson et al., 2022, Fig. 53. Eye area (mm2) (A), facet number (B), and mean facet diameter (μm) (C) for pale and dark colored species of Veromessor. Two species are pale (V. lariversi, V. pseudolariversi—open symbols and regular font), while the other eight species are dark (filled symbols and bold font). For each species, number of workers examined and number of colonies they were derived from is given in parentheses. Significant differences (P < 0.05) among species are denoted after each species name by the letters a–g: a > b > c > d > e > f > g; the three sets of letters for each species correspond to panels A, B, and C, respectively. Groupings are based on univariate F tests within MANCOVA followed by pairwise comparisons using a least significant differences test (see also Johnson & Rutowki, 2022).
,_Fig._53.jpg)
,_Fig._56.jpg)
Keys including this Species
Distribution
Johnson et al. (2022) - The geographic range of V. chamberlini appears to be restricted to coastal islands and adjacent mainland coastal areas of southern California and extending inland along the southern base of the San Gabriel Mountains. Most potential habitat south of the San Gabriel Mountains has been developed. There also is one disjunct record from Santa Clara County that did not indicate a collector, locale, or date. The veracity of this record is unknown, but it seems likely that the historical range of V. chamberlini extended along coastal areas north to or near the San Francisco Bay area. Efforts should be made to reverify this northern record. This species occurs at elevations from 30–1,500 m in the California interior chaparral and woodlands, California coastal sage and chaparral, and California montane chaparral and woodlands ecoregions, as defined by Olson et al. (2001).
 Johnson et al., 2022, Fig. 10. Geographic distribution of: (A) Veromessor andrei, (B) V. chamberlini, and (C) V. chicoensis. The larger black circle in each panel denotes the type locality. The northernmost locale for V. chamberlini was given only as Santa Clara County, and we have placed this locale near the center of the county.
Johnson et al., 2022, Fig. 10. Geographic distribution of: (A) Veromessor andrei, (B) V. chamberlini, and (C) V. chicoensis. The larger black circle in each panel denotes the type locality. The northernmost locale for V. chamberlini was given only as Santa Clara County, and we have placed this locale near the center of the county.
,_Fig._10.jpg)
{kind=link}
Latitudinal Distribution Pattern
Latitudinal Range: 37.213544° to 34°.
| North Temperate |
North Subtropical |
Tropical | South Subtropical |
South Temperate |
- Source: AntMaps
Distribution based on Regional Taxon Lists
Nearctic Region: United States (type locality).
Distribution based on AntMaps
Distribution based on AntWeb specimens
Check data from AntWeb
Countries Occupied
| Number of countries occupied by this species based on AntWiki Regional Taxon Lists. In general, fewer countries occupied indicates a narrower range, while more countries indicates a more widespread species. |

|
Estimated Abundance
| Relative abundance based on number of AntMaps records per species (this species within the purple bar). Fewer records (to the left) indicates a less abundant/encountered species while more records (to the right) indicates more abundant/encountered species. |

|
Biology
Gland chemistry has not been examined in V. chamberlini. Like other small-colony congeners, workers of V. chamberlini have a small pygidial gland reservoir and lack a textured tergal cuticle (Hölldobler et al., 2013; Johnson et al., 2022).
Mating flights have not been observed, but sexuals have been collected in Malaise traps from July 1 through Aug 2, indicating that mating flights occur during summer (Johnson et al., 2022).
Castes
Nomenclature
The following information is derived from Barry Bolton's Online Catalogue of the Ants of the World.
- chamberlini. Messor chamberlini Wheeler, W.M. 1915b: 410 (w.) U.S.A. (California).
- Type-material: 18 syntype workers.
- Type-locality: U.S.A.: California, Santa Cruz I. (R.V. Chamberlin).
- Type-depositories: LACM, MCZC.
- Wheeler, G.C. & Wheeler, J. 1972b: 240 (l.).
- Combination in Novomessor (Veromessor): Emery, 1921f: 67;
- combination in Messor: Bolton, 1982: 341 (in text);
- combination in Veromessor: Wheeler, W.M. & Creighton, 1934: 366; Ward, et al. 2015: 73.
- Subspecies of andrei: Enzmann, J. 1947b: 152 (in key).
- Status as species: Emery, 1921f: 67; Wheeler, W.M. & Creighton, 1934: 366 (redescription); Wheeler, W.M. 1935g: 17; Creighton, 1950a: 159; Smith, M.R. 1951a: 799; Smith, M.R. 1956a: 37 (in key); Smith, M.R. 1958c: 119; Smith, D.R. 1979: 1364; Wheeler, G.C. & Wheeler, J. 1986g: 38 (in key); Bolton, 1995b: 253; Ward, 2005: 66.
- Distribution: U.S.A.
Type Material
- Lectotype worker ((designated by Johnson et al., 2022: 27) from Santa Cruz Island, Santa Barbara County, United States, no date (Dr. R.V. Chamberlin) [USNMENT00529080] [USNM].
- Paralectotypes: 3 workers [LACM], 6 workers [MCZC], 2 workers [USNM], UNITED STATES, California, Santa Barbara County, Santa Cruz Island, no date (Dr. R.V. Chamberlin).
Unless otherwise noted the text for the remainder of this section is reported from the publication that includes the original description.
Description
Worker
Wheeler and Creighton (1934) - Length 4 mm.
Head, exclusive of the mandibles, as broad as long, the sides very slightly narrowed from a point just posterior to the eyes to the insertion of the mandibles. Occiput flat, its angles very broadly rounded. Anterior border of the clypeus virtually straight, the median lobe very shallowly impressed in front and with two feeble carinae. Mandibles large, their external border strongly curved; masticatory margin with two sharp, prominent, terminal teeth and a row of denticles along the rest of the margin. Frontal carinae prominent, moderately divergent anteriorly and strongly divergent behind. The antennal scapes in :repose fail to reach the occipital border by a distance less than their greatest thickness; the proximal end of the scape flattened and forming a spatulate portion which is wider than the feebly clavate tip. First funicular joint slightly shorter than the following two together; funicular joints 2-7 all longer than broad though gradually increasing in thickness, the remaining four joints much more thickened and forming a distinct club. Eyes rather small, feebly convex, suboval, situated at the middle of the side of the head.
Promesonotum in profile strongly though somewhat irregularly convex, with the mesonotum descending abruptly to the much lower epinotum, the mesoepinotal suture feebly impressed. Dorsum of the epinotum very feebly sinuate and much longer than the declivous face, the angle between them armed with two long, feebly arcuate spines which are thin toward the tips but rather thick toward the base. Seen from above, the promesonotum is sub pyriform and notably wider than the remainder of the thorax, with the promesonotal suture obsolescent but rather clearly visible in certain lights. Sides of the thorax not constricted at the mesoepinotal suture, the epinotal spines only moderately divergent and approximately as long as the distance between their tips. Petiole in profile with a long anterior peduncle, which lacks a ventral tooth and increases slightly in diameter toward the node. The sloping anterior face of the node meets the peduncle in a very wide angle, the summit of the node is rounded, the posterior face short and abrupt. The posterior peduncle is short and forms a distinct angle at its junction with the node. Postpetiole in profile with a bluntly angular dorsum and a sinuate ventral face. Seen from above the peduncle of the petiole is compressed anteriorly by two semicircular constrictions, posterior to which it gradually increases in width. Postpetiole from above subpyriform only slightly less than twice as wide as the node of the petiole. Gaster small, oval. Head entirely covered with fine rugae which diverge from the median line toward the occipital angles; interrugal spaces feebly granulose and somewhat shining. Mandibles with coarse longitudinal striae, feebly shining. Clypeus granulose, opaque except for the median lobe which is rather strongly shining. Frontal area opaque. Antennal scapes smooth, strongly shining. Thorax, except for the pronotum where the rugae are transversely arcuate, entirely covered with strong irregular longitudinal rugae, somewhat more shining than the head. Petiolar nodes very feebly granulose, somewhat shining. Gaster very smooth and shining, the small piligerous punctures scarcely visible. Hairs very irregular both in length and thickness. Gular ammo chaetae very poorly developed, consisting of only a few, fine, long hairs. Clypeus with a fringe of rather stout, golden hairs of irregular length. Front, vertex, and occiput with a number of short, fine, whitish hairs many of which are suberect. Thoracic hairs abundant over the entire dorsum and very irregular in length. Petiole, postpetiole, and gaster with abundant whitish hairs which are more uniform in length than those of the thorax. Femora with a few short erect hairs, those of the tibiae, tarsi, and antennal scapes finer, more numerous, and sub erect. Head and thorax dear, ferruginous red, nodes of the pedicel a duller red, base of the gaster yellowish red, femora, tibiae, and posterior gastric segments piceous brown.
References
- Bolton, B. 1982. Afrotropical species of the myrmecine ant genera Cardiocondyla, Leptothorax, Melissotarsus, Messor and Cataulacus (Formicidae). Bulletin of the British Museum (Natural History). Entomology, 46: 307-370 (page 341, Combination in Messor)
- Creighton, W. S. 1950a. The ants of North America. Bulletin of the Museum of Comparative Zoology 104: 1-585 (page 159, Revived status as species)
- Enzmann, J. 1947b. New forms of Aphaenogaster and Novomessor. J. N. Y. Entomol. Soc. 55: 147-152 (page 152, Subspecies of andrei)
- Johnson, R.A., Borowiec, M.L., Snelling, R.R., Cole, A.C. 2022. A taxonomic revision and a review of the biology of the North American seed-harvester ant genus Veromessor (Hymenoptera: Formicidae: Myrmicinae). Zootaxa 52061, 1-115 (doi:10.11646/zootaxa.5206.1.1).
- Plowes, N.J.R., Johnson, R.A., Holldobler, B. 2013. Foraging behavior in the ant genus Messor (Hymenoptera: Formicidae: Myrmicinae). Myrmecological News 18, 33-49.
- Ward, P.S., Brady, S.G., Fisher, B.L. & Schultz, T.R. 2014. The evolution of myrmicine ants: phylogeny and biogeography of a hyperdiverse ant clade (Hymenoptera: Formicidae). Systematic Entomology, DOI: 10.1111/syen.12090
- Wheeler, G. C.; Wheeler, J. 1972b. Ant larvae of the subfamily Myrmicinae: second supplement on the tribes Myrmicini and Pheidolini. J. Ga. Entomol. Soc. 7: 233-246 (page 240, larva described)
- Wheeler, W. M. 1915b. Some additions to the North American ant-fauna. Bull. Am. Mus. Nat. Hist. 34: 389-421 (page 410, worker described)
- Wheeler, W. M.; Creighton, W. S. 1934. A study of the ant genera Novomessor and Veromessor. Proc. Am. Acad. Arts Sci. 69: 341-387 (page 366, Combination in Veromessor)
References based on Global Ant Biodiversity Informatics
- Adams T. A., W. J. Staubus, and W. M. Meyer. 2018. Fire impacts on ant assemblages in California sage scrub. Southwestern Entomologist 43(2): 323-334.
- Johnson R. Personnal Database. Accessed on February 5th 2014 at http://www.asu.edu/clas/sirgtools/resources.htm
- Mallis A. 1941. A list of the ants of California with notes on their habits and distribution. Bulletin of the Southern California Academy of Sciences 40: 61-100.
- Smith M. R. 1956. A key to the workers of Veromessor Forel of the United States and the description of a new subspecies (Hymenoptera, Formicidae). Pan-Pacific Entomologist 32: 36-38.
- Staubus W. J., E. S. Boyd, T. A. Adams, D. M. Spear, M. M. Dipman, W. M. Meyer III. 2015. Ant communities in native sage scrub, non-native grassland, and suburban habitats in Los Angeles County, USA: conservation implications. Journal of Insect Conservervation 19:669–680
- Wetterer, J. K.; Ward, P. S.; Wetterer, A. L.; Longino, J. T.; Trager, J. C.; Miller, S. E. 2000. Ants (Hymenoptera: Formicidae) of Santa Cruz Island, California. Bulletin of the Southern California Academy of Sciences 99:25-31.
- Wetterer, J.K., P.S. Ward, A.L. Wetterer, J.T. Longino, J.C. Trager and S.E. Miller. 2000. Ants (Hymenoptera:Formicidae) of Santa Cruz Island, California. Bulletin of the Southern California Academy of Science 99(1):25-31.
- Wheeler W. M. 1915. Some additions to the North American ant-fauna. Bull. Am. Mus. Nat. Hist. 34: 389-421.
- Wheeler W. M., and W. S. Creighton. 1934. A study of the ant genera Novomessor and Veromessor. Proceedings of the American Academy of Arts and Sciences 69: 341-387.
- Wheeler W.M. 1935. Check list of the ants of Oceania. Occasional Papers of the Bernice Pauahi Bishop Museum 11(11):1-56.